

How to Cite | Publication History | PlumX Article Matrix
Mukhammad Asyari1, Pingkan Aditiawati2, Akhmaloka1, Rukman Hertadi1*
1Biochemistry Research Group, Faculty of Mathematics and Natural Sciences, Institut Teknologi Bandung, Ganesha no. 10 Bandung 40132, Indonesia. 2Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Jl. Ganesha no. 10 Bandung 40132, Indonesia.
DOI : http://dx.doi.org/10.13005/bbra/1856

ABSTRACT: Five lipase genes (lipab4, lipab8, lipab15, lipag18, and lipab18) were isolated from Halomonas and Chromohalobacter local strains of Bledug Kuwu isolates. Based on amino acid sequence analysis, the genes showed some unique motif of amino acid sequences. All of lipases were classified as member of family IV (HSL=hormone-sensitive lipase). These lipases show high similarities of conserved regions with lipolytic of Halomonas and formed a distinct cluster with other types of HSL, such as esterase/lipase and carboxylesterase. All of lipases contain more negative charged of amino acid residues compared to the mesophilic and thermophilic ones, and tend to have similarity to lipases of moderate halophilics. The result of homology and phylogenetic analysis showed that these lipases were clustered into three groups. Group I (lipab8, lipab18 and lipag18) are close to lipolytic gene of Halomonas elongata DSM 2581, while groups II (lipab4) and III (lipab15) create new branches in the phylogenetic tree. In addition, analysis of GC, GC-AT and GC-AT content on the codon usage of the genes revealed the unique profile compared to that the other lipase genes.
KEYWORDS: Halomonas; Chromohalobacter; Halostable lipase; HSL Family; GC-AT content; Bledug Kuwu
Download this article as:| Copy the following to cite this article: Asyari M, Aditiawati P, Akhmaloka, Hertadi R. Cloning and Sequence Analysis of Lipase Gene of Halophilic Bacteria Isolated from Mud Crater of Bledug Kuwu, Central Java, Indonesia. Biosci Biotech Res Asia 2015;12(3) |
| Copy the following to cite this URL: Asyari M, Aditiawati P, Akhmaloka, Hertadi R. Cloning and Sequence Analysis of Lipase Gene of Halophilic Bacteria Isolated from Mud Crater of Bledug Kuwu, Central Java, Indonesia. Biosci Biotech Res Asia 2015;12(3). Available from: https://www.biotech-asia.org/?p=2348 |
Introduction
A lipase is an enzyme that catalyzes hydrolysis of triacylglycerol into fatty acids and glycerol at water-lipid interface, and also an esterification reaction in non-aqueous environment, ie. organic solvents1,2. In some industrial process, the reactions may be inhibited by high concentrations of salt, organic solvents and high temperatures. Therefore, lipases that have optimum activity within wide range of temperatures, salinity, pHs, and various organic solvents, are required by most industries3,4. One of the potential sources of lipases is extremophilic microorganisms, such as thermophiles for thermostable lipase5–10 and halophiles for halostable lipase11–13.
Halophiles are defined as microorganisms that live in hypersaline environments14,15. There are numerous reports about extracellular lipase production from halophiles ie. Salinivibrio sp. strain SA-2, Natronococcus sp., Salicola, Halovibrio, Halomonas, Oceanobacillus, Thalassobacillus, Halobacillus, Virgibacillus, Gracilibacillus, Salinicoccus, and Piscibacillus 16–19 Li and Yu (2012), has been isolated lipase from Chromohalobacter sp. LY7-8, with highly active against p-nitrophenyl esters (C8 to C18) 20. The purified lipase was determined to be 44 kDa, showing optimal activity at 60°C, pH 9.0 and 12.5% NaCl. Significant inhibition of the lipase activity was shown by EDTA, PMSF and DEPC, indicating as metalloenzyme with serine and histidine residues essential for its catalytic function. Perez et al., (2011) has been cloned, overexpressed and purified, a lipolytic enzyme from the moderate halophilic Marinobacter lipolyticus SM19 into E.coli21. The enzyme shows unusual halohilic properties, such as maximal activity in the absence of NaCl and inhibited by presence of NaCl. However the enzyme shows remarkable stability in a wide variety of 30% organic solvents including DMSO, DMF, methanol, ethanol, toluene, diethylether, propanol, and acetone. It has been reported that some halophilic bacteria also live in some organic solvents indicating their enzymes stable and active in those solvents22–24. Isolating lipase from halophilic bacteria, therefore will give higher probability to obtain the enzyme that stable and active in both extreme conditions ie. hypersaline and organic solvents.
Seven halophilic bacteria comprised of four Halomonas, one Chromohalobacter and two Pseudomonas strains were isolated and identified as potential sources for halostable lipases. Four Halomonas and one Chromohalobacter strains showed high halotolerant levels and classified as moderate halophilics, meanwhile the Pseudomonas were classified as low halotolerant level25. One of the Pseudomonas was shown as potential source of organic solvent stable lipase26. Therefore, there are possibility that the other lipase isolated from the same environment is also stable in organic solvents. For further probe the characterized of the lipases, here we present cloning and sequence analysis of the genes.
Materials and Methods
Chemicals
Common chemicals with pro analysis grade were purchased from Merck (Germany) and Sigma-Aldrich (USA). Bacterial growth nutrients, such as tryptone, yeast extract were obtained from Bio Basic (Canada). Biochemical reagents such as dNTPs, PCR Buffer, Taq DNA Polymerase were purchased from Fermentas (USA) and Kapa Biosystems (USA). Oligonucleotides (primers) were ordered from Macrogen (South Korea) and Integrated DNA Technologies (Singapore). Purification of PCR product used GeneJET Gel Extraction Kit (Thermo Scientific). The cloning was performed by using pGEM-T easy vector and T4 DNA ligase that were purchased from Promega (USA). Restriction enzymes were purchased from Thermo Scientific (USA).
Bacteria
Five isolates of halophilic bacteria were obtained from our culture collections in the Laboratory of Biochemistry, Faculty of Mathematics and Natural Sciences, Institut Teknologi Bandung. The isolates were Halomonas meridiana BK-AB4 (AB4), Halomonas elongata BK-AB8 (AB8), Halomonas eurihalina BK-AB15 (AB15), Halomonas elongata BK-AG18 (AG18) and Chromohalobacter japonicus BK-AB18 (AB18)25. Escherichia coli TOP 10 was used as host for gene cloning.
Cultivation
Five isolates were cultivated using modified media of Luria Berthani composed of 0.1% tryptone, 0.05% yeast extract and 10% NaCl27. The cultures were incubated in shaker incubator at room temperature with aeration rate of 150 rpm.
Isolation of Chromosomal DNA
Isolation of chromosomal DNA was carried out using modified method of Zhou28. The collected DNA pellet was separated from the supernatant and dried with concentrator, subsequently followed by resuspension with 25-50 mL ddH2O and stored at 4 oC. The DNA was used for PCR.
Cloning of Lipase Genes
Cloning of lipase gene in each bacterial isolates was started by in vitro amplification of the gene by PCR technique29 using a pair of specific primers ie. xFLipH3A (5’-ATGCAGATCGATGCCTTTCGTCGC-3’) and xRLipH3A (5’-TCATTCCACTCGTCGACCCAGC-3’). A typical PCR mixture (50 µL in volume) was prepared by mixing 5 µL of PCR buffer 10´, 2.5 mM MgCl2, 250 µM of deoxynucleosid-e triphosphate, 0.25 µM of each primers, and 1.25 U of Taq DNA polymerase. PCR was conducted by the following protocol: an initial denaturation was set at 94 oC for 4 min followed by 35 cycles of denaturation (@30 s at 94 oC), an annealing was programmed at 59 oC for 30 s, while an extension and final extension were carried out at the same temperature at 72 oC for 1 min and 5 min, respectively29. The product of PCR was verified by electrophoresis technique using submerged horizontal electrophoresis cell for 50 min at 70 volts. The purification of PCR product was carried out by GeneJET Gel Extraction Kit. Finally, purified DNA was re-suspended to 50 µL buffers (10 mM Tris-HCl, pH 8.5). The purified DNA solution was stored at -20 oC.
Construction of recombinant plasmid was carried out by ligating the PCR products with pGEM-T easy plasmid. The competent cell of E. coli Top 10 was prepared following the method described by Cohen30. Transformation of E. coli was conducted by heat shock method31. 100 mL of the transformed cells were spreaded on LB agar (LBA) media containing 100 mg/mL of ampicillin, 15 mg/mL of tetracycline and incubated at 37 oC for overnight. Plasmid isolation from the transformed cell was carried out by the alkaline lysis method32. Finally to verify the recombinant plasmid containing DNA insert, the plasmid was digested with EcoRI and visualized by agarose gel electrophoresis.
Sequencing of Lipase Gene
Lipase genes were sequenced by Dideoxy-Sanger dye terminator method33 at Macrogen, Seoul, Republic of Korea. The sequences were validated by analyzing the electrophoregram data using Sequence scanner 2 (Applied Biosystems). In order to combine the partial gene sequences into a full gene, we used DNA Baser Sequence Assembler v3 program (Heracle BioSoft).
Deduced Amino Acid Sequence Analysis of Lipase
Deduced amino acid sequences of five lipases were performed by in silico translation using Bioedit software and online server of ExPASy-Translate tool available at http://web.expasy.org/translate/.
Homology analysis of five lipase sequences were carried out using NCBI-Blastp analysis program (http://blast.ncbi.nlm.nih.gov/ Blast.cgi). A hundred of high homologous sequences were used to generate phylogenetic profile using MEGA 6 software based on the Neighbor-Joining clustering method.
Amino acid composition and alignment analysis were performed using “Amino Acid Composition” and “ClustalW Multiple Alignment” programs which are integrated in the Bioedit software.
Sequence Analysis of Lipase Gene
The further analysis of lipase gene sequences was carried out by evaluating contents of GC, GC-AT, and GC-AT of codon usage. The last two analyses were conducted with Bioedit-software, while the third analysis was calculated with an online program available at http://www.bioinformatics.org /sms2/ codon_usage.html. The analysis of “codon usage” was performed on the 1st, 2nd and 3rd base position. All analyzes above were not only performed on the five isolates lipase sequences but also to other lipase genes from the other microorganism.
Results and Discussion
Cloning and Sequencing of Lipase Genes
Five lipase genes have been successfully amplified from Halomonas and Chromoholobacter local strains by PCR technique. The genes, namely lipab4, lipab8, lipab15, lipag18, and lipab18, have estimated length about 800 bp (Figure 1). All of the genes were cloned into E. coli Top 10, sequenced and deposited to GenBank database (https://www.ncbi. nlm.nih.gov) with accession number of KJ676135.1, KJ676136.1, KJ676137.1, KJ676139.1, and KJ676138.1, respectively.
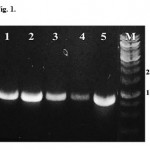 |
Figure 1: The results of PCR amplifications of lipase genes. Assigned lane number is as follow 1=lipab4, 2= lipab8, 3= lipab15, 4= lipab18, 5=lipag18 and M=DNA marker. |
Homology and Phylogenetic Analysis of Lipases
Homology analysis of deduced amino acid sequences of five lipases were carried out using NCBI-Blastp program. The results showed that five lipase sequences showed high similarities to lipase of Halomonas elongata DSM 258134 with percent identity of 95-99%, Gammaproteobacteria MFB02135 with percent identity of 58-60% and Halomonas halodenitrificans36 with percent identity of 57%. All lipases were closed to Halomonas elongata DSM 2581 (Table 1).
 |
Table 1: The result of Blastp alignment for five lipase genes |
Phylogenetic analysis generated a tree profile that five lipases distributed into three groups ie. group I (lipab8, lipab18 and lipag18), groups II (lipab4) and III (lipab15). Group I are closed to lipase of Halomonas elongata DSM 2581, meanwhile the group II and III formed new branches (Figure 2).
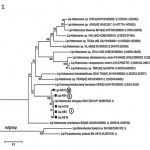 |
Figure 2: Phylogenetic profile generated based on lipase sequences of the five isolates |
Conserved Regions Analysis of The Lipases
Alignment of 100 sequences best homolog to the isolates showed that a few regions are conserved, such as GGGX type, GXSXG (pentapeptide), oxyanion and catalytic triad ie. serine, aspartate and histidine (Figure 3). The presence of these conserved regions suggesting that the isolates classified as member of family IV (HSL=Hormone-Sensitive Lipase). Previous study showed that there are a number of bacterial proteins similar with mammalian HSL37. Moreover, family members of HSL are very wide, including lipases, esterases and carboxylesterases2. Determination of HSL sub family of the lipases were performed based on similarities profiles of conserved regions compared to the lipolytic, lipases, esterases/lipases, esterases and carboxylesterases on the of HSL family members. The results showed that there are few different regions among the isolates compared to that other lipases, as indicated on numbers 1-15 (Figure 4). The lipases are more similar to lipolytic of Halomonas. Therefore, suggesting the lipases are members of moderate halophilic group.
 |
Figure 3: Conserved region of lipase includes GGGX, GXSXG (pentapeptide), oxyanion and catalytic triad were contained in 100 lipase sequences which has closely related to lipase of 5 isolates. |
 |
Figure 4: Differences of conserved region among sequences of lipolytic, lipase, esterase/lipase, carboxylesterase that have close relations with lipase from five isolates. |
Detail analysis by comparing the composition of amino acid residues showed that the amino acid composition of the lipases are more negative charged residues compared to that the mesophilic and thermophilic (Figure 5). However, the negative charged residues of five lipases are close to moderate halophilic than extreme halophilic. The negative charged residues of an extreme halophilic lipases are usually used to stabilize the structure due to the influence of high salt concentrations38.
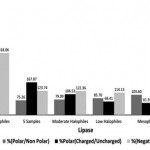 |
Figure 5: Amino acid difference of lipase sequence between five isolates with other related microorganisms. |
Further analysis on the lipases by comparing the amino acid sequences among the five isolates with Halomonas elongata DSM 2581 showed that there are few amino acid differences at several positions. The differences were occurred at position of 98CàR in lipab15 and lipag18, 168QàE in lipab4 and lipab15, 188AàD in lipab4, 191DàG and 197DàG in lipab15 and 199DàA in lipab4, lipab8, lipab15 and lipab18 (Table 2). However, all of the differences do not occur in conserved regions, such as the catalytic site, pentapeptide or oxyanion regions.
 |
Table 2: Amino acid differences between lipase of five isolates with the Halomonas elongata |
Content of GC, GC-AT And GC-AT Codon Usage of Lipase Genes
Previous studies by comparing GC content or GC content on the codon usage, showed a unique characteristic of halophilic. The high GC content of the genome is a common feature of extreme halophilic39,40,41. GC content on third (wobble) position of codon usage is corresponding to over-representation of the translated amino acid, which is for halophilic proteins correspond tp the abundance of acidic residues39. Therefore, overrepresentation of acidic residues is an adaptation to high salinity that is apparent in the codon usage of the organism.
Based on the above studies, we were further investigated the halophilic properties by comparing content of GC, GC-AT and GC-AT of codon usage of the lipase genes with other microorganism. GC content analysis showed that the five lipase genes are closer to all groups of halophilic (Figure 6). The similarities of GC content of the genes with those of halophilic are unique characteristics of the isolates. This was due to corelate to their niches at high salinity level. Meanwhile, the result of GC-AT content analysis of the genes further exhibited a unique pattern as follows, 5 samples = average of halophiles > thermophiles > mesophiles (Figure 7). Further investigation, we analyzed GC-AT content of codon usage of first, second and third positions. The result showed that the profile of GC-AT content for the third base displayed similar unique pattern to the last two analysis (Figure 8). GC-AT content for the third base of codon usage of the genes may be used as differentiator of halophilic lipase from various groups of microorganism. In general, the third base position of the gene is determinant base of codon preferences for each microorganism42. All of the analysis above suggest that the lipase genes of five isolates are closer to moderate halophilic, which is Halomonas elongata DSM 2581.
 |
Figure 6: Percentage GC content of lipase genes of five isolates and other related microorganism |
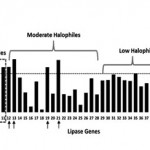 |
Figure 7: GC-AT content of lipase between five isolates and the other related microorganism. Identity of each numbered microorganism is as follow: |
 |
Figure 8: The difference of bases GC-AT content fractions of “usage codon” of the lipase genes between five isolates with the other related microorganism. Two moderates halophilics have similar GC-AT content of 1st, 2nd and 3rd usage codon pattern compared to five lipase genes ie. (12). Halomonas elongata DSM 2581, (13). |
Conclusion
Lipase genes were isolated from five moderately halophilic bacteria of Bledug Kuwu (BK) isolates and cloned into E. coli Top 10. The significant differences of amino acids lipase of five isolates occurs in several positions and exhibit pattern of conserved regions. The lipase of five isolates is classified as a member of family IV lipase (HSL=Hormone-Sensitive lipase) as indicated by the existence of Gly-X-Ser-X-Gly pattern in their conserved regions. The five lipases have high similarities of conserved regions with lipolytic Halomonas and formed a distinct cluster with other types of HSL lipase, such as esterase/lipase and carboxylesterase. Amino acids composition of five lipases are more similar to moderate halophilic, they have more negative charged than mesophilic and thermophilic ones. Homology and phylogenetic profile of five lipase genes distributed them into three groups i.e. Group I (lipab8, lipab18 and lipag18) are closely related to lipolytic gene of Halomonas elongata DSM 2581, while groups II (lipab4) and III (lipab15) create new branches in the phylogenetic tree. Narrowing analysis to GC and GC-AT content have successfully revealed the unique pattern of five lipase genes compared to other microorganism, such as thermophiles and mesophiles. More specific analysis on GC-AT content of codon usage exhibited consistent result with both morphology and phylogenetic analyses above.
Acknowledgements
This work is funded by university priority research grant with the contract number: 310u/11.C01/PL/2015 awarded to R.H.
References
- Gill J, Parish JH. Minireview: Lipases—enzymes at an interface. Biochem Educ. 1997;25(1):2-5.
- Arpigny JL, Jaeger K-E. Bacterial lipolytic enzymes: classification and properties. Biochem J. 1999;343(1):177-183.
- Hasan F, Shah AA, Hameed A. Industrial applications of microbial lipases. Enzyme Microb Technol. 2006;39(2):235-251.
- Ghasemi Y, Rasoul-Amini S, Kazemi a., Zarrini G, Morowvat MH, Kargar M. Isolation and characterization of some moderately halophilic bacteria with lipase activity. Microbiology. 2011;80(4):483-487.
- Widhiastuty MP, Febriani, Moeis MR, Akhmaloka, Madayanti F. Cloning, homological analysis and expression of Lipase gene from hot spring isolate. Int J Integr Biol. 2011;11(1):8-13.
- Febriani, Ihsanawat, Hertadi R, Madayanti F, Akhmaloka. Thermostable alkaline lipase isolated from thermus aquaticus. Int J Integr Biol. 2013;14(2):104-112.
- Aditiawati P, Yohandini H, Madayanti F, Akhmaloka. Microbial diversity of acidic hot spring (kawah hujan B) in geothermal field of kamojang area, west java-indonesia. Open Microbiol J. 2009;3:58-66.
- Akhmaloka A, Suharto a., Nurbaiti S, N. Tika I, M. Warganegara F. Ribotyping Identification of Thermophilic Bacterium from Papandayan Crater. ITB J Eng Sci. 2006;38(1):1-10.
- Nurhasanah, Nurbaiti S, Warganegara FM, Akhmaloka. Diversity of gene encoding thermostable Lipase from compost based on metagenome analysis Isolation of total DNA community from compost. Int J Integr Biol. 2015;16(1):7-12.
- Brilliantoro R, Zidny R, Widhiastuty MP, Akhmaloka. Hydrolytic and Transesterification Activities of Thermostable Lipase ITB1.1. Biosci Biotechnol Res Asia. 2015;12(April):1-6.
- Gupta GN, Srivastava S, Khare SK, Prakash V. Extremophiles: An Overview of Microorganism from Extreme Environment. Int J Agric Environ Biotechnol. 2014;7(2):371-379.
- Sánchez-Porro C, Martín S, Mellado E, Ventosa a. Diversity of moderately halophilic bacteria producing extracellular hydrolytic enzymes. J Appl Microbiol. 2003;94(2):295-300.
- Kumar S, Karan R, Kapoor S, Singh SP, Khare SK. Screening and isolation of halophilic bacteria producing industrially important enzymes. Brazilian J Microbiol. 2012;43(4):1595-1603.
- Litchfield CD. Survival strategies for microorganisms in hypersaline environments and their relevance to life on early Mars. Meteorit Planet Sci. 1998;33(4):813-819.
- Ma Y, Galinski E a., Grant WD, Oren A, Ventosa A. Halophiles 2010: Life in saline environments. Appl Environ Microbiol. 2010;76(21):6971-6981.
- Amoozegar MA, Salehghamari E, Khajeh K, Kabiri M, Naddaf S. Production of an extracellular thermohalophilic lipase from a moderately halophilic bacterium, Salinivibrio sp. strain SA-2. J Basic Microbiol. 2008;48(3):160-167.
- Boutaiba S, Bhatnagar T, Hacene H, Mitchell D a., Baratti JC. Preliminary characterisation of a lipolytic activity from an extremely halophilic archaeon, Natronococcus sp. J Mol Catal B Enzym. 2006;41(1-2):21-26.
- Ozcan B, Ozyilmaz G, Cokmus C, Caliskan M. Characterization of extracellular esterase and lipase activities from five halophilic archaeal strains. J Ind Microbiol Biotechnol. 2009;36(1):105-110.
- Rohban R, Amoozegar MA, Ventosa a. Screening and isolation of halophilic bacteria producing extracellular hydrolyses from Howz Soltan Lake, Iran. J Ind Microbiol Biotechnol. 2009;36(3):333-340.
- Li X, Yu H-Y. Characterization of a novel extracellular lipase from a halophilic isolate, Chromohalobacter sp. LY7-8. African J Microbiol Res. 2012;6(14):3516-3522.
- Pérez D, Martín S, Fernández-Lorente G, et al. A novel halophilic lipase, LipBL, showing high efficiency in the production of eicosapentaenoic acid (EPA). PLoS One. 2011;6(8):1-11.
- Tiquia SM, Davis D, Hadid H, et al. Halophilic and halotolerant bacteria from river waters and shallow groundwater along the Rouge River of southeastern Michigan. Environ Technol. 2007;28(3):297-307.
- Sellek G a., Chaudhuri JB. Biocatalysis in organic media using enzymes from extremophiles. Enzyme Microb Technol. 1999;25(6):471-482.
- Doukyu N, Ogino H. Organic solvent-tolerant enzymes. Biochem Eng J. 2010;48(3):270-282.
- Asy’ari M, Parwata IP, Aditiawati P, Akhmaloka, Hertadi R. Isolation and Identification of Halostable Lipase Producing Bacteria from the Bledug Kuwu Mud Crater Located at Purwodadi-Grobogan , Central Java , Indonesia. J Pure Appl Microbiol. 2014;8(5):3387-3396.
- Parwata IP, Asyari M, Hertadi R. Organic Solvent-Stable Lipase from Moderate Halophilic Bacteria Pseudomonas stutzeri Isolated from the Mud Crater of Bleduk Kuwu , Central Java , Indonesia. J Pure Appl Microbiol. 2014;8(1):31-40.
- Sezonov G, Joseleau-Petit D, D’Ari R. Escherichia coli physiology in Luria-Bertani broth. J Bacteriol. 2007;189(23):8746-8749.
- Stach JE., Bathe S, Clapp JP, Burns RG. PCR-SSCP comparison of 16S rDNA sequence diversity in soil DNA obtained using different isolation and purification methods. FEMS Microbiol Ecol. 2001;36(2-3):139-151.
- Mullis K, Faloona F, Scharf S, Saiki R, Horn G, Erlich H. Specific Enzymatic Amplification of DNA In Vitro: The Polymerase Chain Reaction. Cold Spring Harb Symp Quant Biol. 1986;51:263-273.
- Cohen SN, Chang AC, Hsu L. Nonchromosomal antibiotic resistance in bacteria: genetic transformation of Escherichia coli by R-factor DNA. Proc Natl Acad Sci U S A. 1972;69(8):2110-2114.
- Inoue H, Nojima H, Okayama H. High efficiency transformation of Escherichia coli with plasmids. Gene. 1990;96(1):23-28.
- Bimboim HC, Doly J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979;7(6):1513-1523.
- Sanger F, Nicklen S, Coulson a R. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci U S A. 1977;74(12):5463-5467.
- Schwibbert K, Marin-Sanguino A, Bagyan I, et al. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581 T. Environ Microbiol. 2011;13(8):1973-1994.
- Joseph TC, Baby A, Reghunathan D, Varghese AM, Murugadas V, Lalitha K V. Draft Genome Sequence of the Halophilic and Highly Halotolerant Gammaproteobacteria Strain MFB021. Genome Announc. 2014;2(6):e01156-14 – e01156-14.
- Sakurai T, Nakashima S, Kataoka K, Seo D, Sakurai N. Diverse NO reduction by Halomonas halodenitrificans nitric oxide reductase. Biochem Biophys Res Commun. 2005;333(2):483-487.
- Hemilä H, Koivula TT, Palva I. Hormone-sensitive lipase is closely related to several bacterial proteins, and distantly related to acetylcholinesterase and lipoprotein lipase: identification of a superfamily of esterases and lipases. Biochim Biophys Acta. 1994;1210(2):249-253.
- Lanyi JK. Salt-dependent properties of proteins from extremely halophilic bacteria. Bacteriol Rev. 1974;38(3):272-290.
- Kennedy SP, Ng WV, Salzberg SL, Hood L, DasSarma S. Understanding the adaptation of Halobacterium species NRC-1 to its extreme environment through computational analysis of its genome sequence. Genome Res. 2001;11(10):1641-1650.
- Paul S, Bag SK, Das S, Harvill ET, Dutta C. Molecular signature of hypersaline adaptation: insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008;9(4):R70.1-R70.19.
- Soppa J. From genomes to function: Haloarchaea as model organisms. Microbiology. 2006;152(3):585-590.
- Andersson SG, Kurland CG. Codon preferences in free-living microorganisms. Microbiol Rev. 1990;54(2):198-210.

This work is licensed under a Creative Commons Attribution 4.0 International License.



