

How to Cite | Publication History | PlumX Article Matrix
Screening Rice (Oryza Sativa L.) Genetic Resources for Cold Tolerance at the Germination Stage
Zhanbyrbayev Y. A.1,2, Rysbekova A. B.2,3, Usenbekov B. N.2, Kazkeev D. T.2, Sartbayeva I. A.2, Berkimbay Kh. A.2, Sarsenbayev B. A.2 and Zelensky G. L.4
1Kazakh National Agrarian University, Almaty, 050010, Kazakhstan.
2Institute of Plant Biology and Biotechnology, Almaty, 050040, Kazakhstan.
3S.Seifullin Kazakh Agrotechnical University, Astana 010011,Kazakhstan.
4Federal State Budgetary Scientific Institution All-Russian Rice Research Institute, 350921, Russia, Krasnodar, p. Belozerniy, 3.
Corresponding Author E-mail: bakdaulet7@yandex.ru
DOI : http://dx.doi.org/10.13005/bbra/2351

ABSTRACT: It was conducted the laboratory screening of local and foreing rice samples total amount of 29 genotypes to cold tolerance and cold sensivity. As a result of screening for the first time the most perspective genotypes (Lazurnyi, UzROS 7-13 and Kuban 3) were selected as donors to create the initial cold tolerance forms for rice breeding in Kazakhstan.
KEYWORDS: foreing rice; perspective genotypes ; rice breeding
Download this article as:| Copy the following to cite this article: Zhanbyrbayev Y. A, Rysbekova A. B, Usenbekov B. N, Kazkeev D. T, Sartbayeva I. A, Berkimbay Kh. A, Sarsenbayev B. A, Zelensky G. L. Screening Rice (Oryza Sativa L.) Genetic Resources for Cold Tolerance at the Germination Stage. Biosci Biotech Res Asia 2016;13(4). |
| Copy the following to cite this URL: Zhanbyrbayev Y. A, Rysbekova A. B, Usenbekov B. N, Kazkeev D. T, Sartbayeva I. A, Berkimbay Kh. A, Sarsenbayev B. A, Zelensky G. L. Screening Rice (Oryza Sativa L.) Genetic Resources for Cold Tolerance at the Germination Stage. Biosci Biotech Res Asia 2016;13(4). Available from: https://www.biotech-asia.org/?p=16701 |
Introduction
The Republic of Kazakhstan is the most northern rice-growing area in the world. The rice-growing areas can be located up to 44º 51́ of north latitude (I zone). In natural features, rice-growing area of the republic can be divided into three climatic zones:
The Republic of Kazakhstan is the most northern rice-growing area in the world. The areas of rice crops can be located up to 44º north latitude 51 (I zone). In natural features rice-growing area of the republic can be divided into three climatic zones: I North – Karatal, Akdala, Tasmurun, Charyn, Kazalinsk areas. Sum of temperatures is (2700-3250 ºC) during the growing season of rice with temperature above 15 °C.
II Central – Kyzylorda, Shieli areas. Sum of temperatures is (3250-3600 ºC) during the growing season of rice with temperature above 15 °C.
III South – Togusken, Kyzylkum areas with a total temperature 3600ºS.
The boundaries of the zones are held by the isolines of average sums of temperatures above +15°C: for 3100, 3300 and 3500 °C, characterizing the optimum conditions for the full annual ripening of the early-ripening, mid-ripening and late-ripening varieties of rice [1].
The total area of the equipped systems for rice cultivation in account of rice crop rotation is about 225 thousands ha. According to statistics, from 280 to 350 thousand tons of raw rice is produced every year in Kazakhstan. 90% of Kazakhstan rice crops are located in the main rice-growing region of the Republic – Kyzylorda region.
Unstable seasonal atmospheric pressure in different years leads to sharp changes in climatic conditions. Therefore, now it is an acute problem in the Republic of Kazakhstan to create varieties resistant to cold stress in the initial period of growth and development of rice plants having a high germination and increased energy of growth at the adverse temperature conditions.
Low temperatures lead to a number of adverse effects: they reduce the growth energy of seeds and photosynthetic activity, cause discoloration of leaves, reduce plant height and productivity of spikelets in the panicles, cause degeneration of spikelets in the panicles, delay of paniculation, uneven ripening of grains and the poor quality of the grain. Effect of low temperatures is revealed in the various stages of growth of rice plants [2]. In this regard, the first and most important step in rice breeding programs by tolerance to cold stress is the initial screening of breeding samples to identify of donors that tolerance to this stress factor. It was found that the cold tolerance of rice is a complex polygenic trait and it is caused by the simultaneous action of a number of physiological and biochemical mechanisms affecting various metabolic processes [3].
It is known that the rice genotypes of subspecies japonica is more resistant to low temperatures and have higher cold tolerance as compared with such traits of subspecies indica. Quantitative indication of cold tolerance of rice at seedling stage is controlled by two genes Cts-1 and Cts-2 (t); in the booting stage – by multiple genes. Although genetic mechanisms of rice cold tolerance at this stage of ontogenesis is not studied sufficiently, however, some of them were identified successfully using QTL analysis. For example, two closely related quantitative loci (Ctb-1 and Ctb-2) of cold tolerance associated with the anther length were identified by Saito et. al.. [4]. Takeuchi et al. created a genetic linkage map using RAPD and RFLP molecular markers and they found in sum eight quantitative loci of cold tolerance [5]. QTL-analysis, conducted by another research group, allowed to identify three QTL-loci (qPSST-3, qPSST-7, and qPSST-9), whose total contribution to the phenotypic variation for a given trait is 27.4%. In addition, the authors identified SSR-markers RM569, RM1377, RM24545, which were linked closely with these loci. The percentage of germinated seeds in the irrigation water at a temperature of 18-19 °C and at controlled temperature (17 °C) in the greenhouse was used as a phenotypic test on cold tolerance [6].
The main aim of this study was to evaluate the cold tolerance of Kazakhstan, Russia and other countries rice breeding materials at the seed germination stage.
Material and Methods.
Twenty nine rice varieties were used in this study, 5 belonging to japonica and 24 to indica subspecies from different ricegrowing countries (Table 1).
Cold tolerance evaluation
According to the Institute of rice, varietal differences in the rate of germination of rice seeds is most clearly manifested at low temperature 14 °C, and these differences are smoothed at lower and higher temperatures. 75 samples of seeds were immersed first into 12% H2O2 for 15 minutes to destroy the harmful microflora in the weevil, and then they were washed 2 times with distilled water. Treated seeds were immersed in water at room temperature for 2 hours to accelerate the swelling, and then they were laid out in the Petri dishes with moistened filter paper (two layers) in 3-fold replicates, with 25 weevil samples in each dish, and were placed in a climatic chamber at a constant temperature of 14 °C, at the photoperiod of 16 hours/day and 8 hours/night and at illumination and 50µE m-2 S-1 klx. Control plants were grown at temperature 28°C. Germination of the part of weevils were observed on the 2nd day in the control plants, and on the 5th day in the experimental one, the amount of germinated seeds was counted. The following counts were carried out every other day. Counting was finished on the 13th day of the experiment, and germination energy of rice seeds was measured [7]. Along with seed germination energy, seeds viability was calculated in the control and experimental samples.
Results
In the laboratory, 29 rice samples were screened for cold tolerance and cold sensitivity. The Russian variety Kuban 3 was used as a standard of the cold tolerance. Criterion for germination energy (GE) evaluation is usually presented not by the percentage of seeds germinated over a certain period, but by “the average duration of germination of one seed”, showing the conventional number of days required for germination of the single seed. If the average germination energy is, for example, 4.8 days for the first sample and the second sample is characterized by a large number of days, this would indicate that the seeds of the second sample have a lower germination energy, since it takes more time for germination of one seed. On the 13th day of the experiment, seeds viability, germination energy and length of coleoptile in the studied genotypes at temperature 14 °C was determined (Table 1).
Table 1: Country of origin, subspecies and germination of 29 rice varieties under 280C (control) and 140C (treatment) growth regimes (data presented are the means of 3 replications)
| Genotype | Country of Origin | Subspecies | Control (28 °С) | Treatment (14 °С) | ||
| Germination, % | GE, days | Germination, % | GE, days | |||
| Avangard | Uzbekistan | japonica | 82.5 | 3.0 | 36.0 | 10.9 |
| Akdala | Kazakhstan | japonica | 100.0 | 3.7 | 44.0 | 9.4 |
| Altynay | Kazakhstan | japonica | 95.0 | 2.8 | 40.0 | 8.1 |
| Anait | Russia | japonica | 90.0 | 2.8 | 52.0 | 9.2 |
| Aral 202 | Kazakhstan | japonica | 100.0 | 2.7 | 56.0 | 9.8 |
| Aru | Kazakhstan | japonica | 92.5 | 3.0 | 40.0 | 9.3 |
| Bakanasski | Kazakhstan | japonica | 100.0 | 2.7 | 52.0 | 9.7 |
| Izumrud | Russia | indica | 100.0 | 2.2 | 20.0 | 9.0 |
| KazNIIR-5 | Kazakhstan | japonica | 95.0 | 2.2 | 80.0 | 8.3 |
| Ко 293 | Philippines | indica | 100.0 | 3.5 | 20.0 | 10.6 |
| Kuban 3 | Russia | japonica | 100.0 | 3.5 | 100.0 | 7.4 |
| Kurchanka | Russia | japonica | 87.5 | 3.1 | 4.0 | 9.4 |
| Lazurnyi | Uzbekistan | indica | 85.0 | 3.1 | 76.0 | 6.4 |
| Lider | Russia | japonica | 82.5 | 3.2 | 72.0 | 8.6 |
| Liman | Russia | japonica | 82.5 | 3.3 | 28.0 | 9.6 |
| Madina | Kazakhstan | japonica | 97.5 | 2.9 | 44.0 | 9.6 |
| Marzhan | Kazakhstan | japonica | 67.5 | 3.7 | 40.0 | 9.5 |
| Novator | Russia | japonica | 75.0 | 2.4 | 68.0 | 9.3 |
| Opytnoe | Kazakhstan | japonica | 87.5 | 3.4 | 36.0 | 8.1 |
| PakLi | Kazakhstan | indica | 80.0 | 2.5 | 28.0 | 10.2 |
| Snezhinka | Russia | japonica | 100.0 | 2.9 | 28.0 | 9.6 |
| Solnechnyi | Russia | japonica | 65.0 | 3.2 | 52.0 | 9.4 |
| Taibonet | Italy | indica | 67.5 | 4.1 | 4.0 | 11.0 |
| UzRos 7-13 | Uzbekistan | japonica | 85.0 | 3.3 | 68.0 | 6.6 |
| Fisht | Russia | japonica | 75.0 | 3.9 | 84.0 | 8.29 |
| Sharm | Russia | japonica | 97.5 | 2.7 | 16.0 | 9.6 |
| Yantar | Russia | japonica | 95.0 | 2.9 | 36.0 | 8.4 |
| Jinbubyeo | Korea | japonica | 70.0 | 2.8 | 54.0 | 10.8 |
| Odaebyeo | Korea | japonica | 75.0 | 2.4 | 49.0 | 10.6 |
In the control variant, about 50% of the samples showed a high level of seed germination (˃95%) already on the 2nd day, while the experimental samples showed the same only on the 5th day. The genotypes were divided into 3 groups by seeds viability: cold-tolerant (T), 66-100% of viability, moderate (M), the germination of 34-66% of viability, and cold sensitive (S), the viability of 0-33%. In cold-tolerant genotypes Lazurnyi, Kuban 3 and UzRos 7-13 seed germination have been started on the 5th day already. Korean standards of cold tolerance (Jinbubyeo, Odaebyeo) and local released varieties, which belong to the subspecies japonica (except KazNIIR 5), such as Opitnoe, Altynay, Aru, Marzhan, Akdala, Madina, Bakanassky and Aral 202, were referred by us as a moderate. Rice varieties Taibonet, Sharm, Izumrud, Ko 293 IRRI, PakLi and Snezhinka are classified as cold sensitive. The obtained data indicate that the tested rice samples differ by germination energy. This rate ranged from 2.2 to 4.1 days in the control samples at temperature 28 °C, regardless to the systematic and geographical origin of varieties. Among the experimental samples at 14 ºC, varieties of breeding of Uzbek Research Institute of rice (Lazurnyi, UzRos 7-13) and the All-Russia Research Institute of rice (Kuban 3) were characterized by the fewest days on seed germination rate. In average, this factor was 6-7 days at varieties Lazurnyi, UzRos 7-13 and Kuban 3 and it gives a reason to include them in the breeding on the creation of initial cold tolerance material.
The samples under research were divided into 4 groups on the basis of the studied trait. The group I includes genotypes with the least GE, germination of such seeds requires the least amount of days (6.4-7.4). Group II, dramatically different from the first, consists of genotypes characterized by average GE (8.1-8.6 days). Groups III and IV include genotypes with high GE: 8.8-9.8 and 10.2-11.0 days, respectively (Figure1).
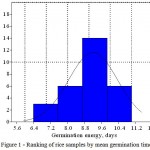 |
Figure 1: Ranking of rice samples by mean germination time
|
It was found that 14 of the 29 studied genotypes can belong to the third group, which gives a reason to consider them as moderate.
According to Cruz and Milach (2004), a clear difference between cold-sensitive and cold-tolerant genotypes can be revealed just by change of the length of the coleoptile [8]. In our experience, three replicates were combined into one, and then 20-30 seedlings were selected by the method of average samples and length of coleoptile was measured in them, and then the general average value of that body was calculated in the test sample. In this study, clear regularity was observed at the measurement of the length of the seedlings coleoptile on the 13th day: the length of coleoptile increases at short term GE. For example, if Lazurnyi, UzROS 7-13 and Kuban 3 were characterized by a short term GE, but by the highest values of the coleoptile length – (0.67 to 0.76 cm) (Figure 2).
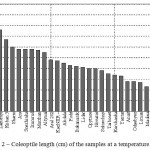 |
Figure 2: Coleoptile length (cm) of the samples at a temperature of 14 0C |
Thus, by screening of 29 genotypes it was revealed that genotypes differ in coleoptile length and they can be divided into three groups by a given trait:
1) short coleoptile – 0.12 – 0.33 mm;
2) medium coleoptile – 0.34 – 0.55 mm;
3) long coleoptile – 0.56 mm or more (Figure 3).
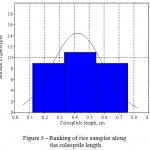 |
Figure 3: Ranking of rice samples along the coleoptile length |
Experimental data suggest that with increasing germination energy in most genotypes the growth rate of coleoptile is reduced. The correlation indicates an inverse relationship between these symptoms (r = -0,60). The revealed laws indicate to the inappropriateness of the inclusion of genotypes Avangard, PakLee, Taibonet into the breeding schemes by cold tolerance.
The research allowed to evaluate rice varieties from different countries breeding by cold tolerance in various stages of ontogeny. With respect to GE, the studied samples were divided into 4 groups. It was established that 14 of the 29 studied genotypes are from the third group, and considered as medium cold-tolerant genotypes.
Among the studied varieties, Taibonet, Sharm, Izumrud, Ko 293, PakLee and Snezhinka, which belong to the subspecies indica, proved to be cold sensitive, and it is rightly so. It is known that subspecies indica are distributed mainly in subtropical areas and advantageously in the tropics. Therefore, varieties of subspecies indica do not demonstrate intensive growth in the early stages because of the greater adaptability to interchange culture and they form very thinned seedlings at direct seeding followed by a complete flooding, and often they die completely. In our studies, cold-tolerant variety Lazurnyi of breeding of Uzbek Research Institute of rice is an exception of the rule, as it also belongs to the subspecies indica. In the result of screening in the early phase of ontogenesis, the rice varieties Kuban 3 (Russian cold tolerant standard), UzRos 7-13 and Lazurnyi proved to be the most cold-tolerant ones among all the samples under research.
In general, it should be noted that the screening researches of the collection samples reveal the donors of agronomic traits of rice (and other crops), which is extremely important to create pre-breeding resources.
References
- Kovalenko V.I., Dudenko V.P. Rice in Kazakhstan // Alma-ata, 1974. 176 p.
- Suh J.P., Jeung J.U., Lee J.I., Choi Y.H., et al. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerance genotypes of rice (Oryza sativa) // Theor. Appl. Genet. 2010, 120: P. 985–995.
CrossRef - Kostylev P.I., Stepovoy V.I., Parfenuk A.A. North rice. Rostov-na-Donu. “Book”, 2004, 576 p.
- Saito, K., Y. H. Saito, W. M. Funatsuki, Y. Sato et al. Physical mapping and putative candidate gene identification of a quantitative trait locus Ctb1 for cold tolerance at the booting stage of rice // Theor. Appl. Genet. 2004, Р. 109:515-522.
- Takeuchi, Y., H. Hayasaka, B. Chiba, I. Tanaka, T. et al. Mapping quantitative trait loci controlling cool-temperature tolerance at booting stage in temperate japonica rice // Breed. Sci. 2001, Р. 51:191–197.
- Suh J.P., Jeung J.U., Lee J.I., Choi Y.H., et al. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerance genotypes of rice (Oryza sativa) // Theor. Appl. Genet. 2010, 120, P. 985–995.
CrossRef - Skazhennik M.A., Vorobiev N.V., Doseeva O.A. Methods of physiological research in rice growing. Krasnodar: Rice Research Institute, 2009. – 23 p.
- Cruz R.P., Milach S.C.K, Federizzi L.C. Inheritance of rice cold tolerance at the germination stage // Genetics and Molecular Biology. 2006, 29, P. 314 – 320.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.



