

How to Cite | Publication History | PlumX Article Matrix
Noble Kiriyachan Kurian1 and Sarita Ganapathy Bhat2
and Sarita Ganapathy Bhat2
1Department of Biotechnology, Cochin University of Science and Technology, Kalamassery, Cochin-22, Kerala, India.
2Department of Biotechnology, Cochin University of Science and Technology, Kalamassery, Cochin-22, Kerala, India.
Corresponding Author E-mail saritagbhat@gmail.com
DOI : http://dx.doi.org/10.13005/bbra/2594

ABSTRACT: Photoprotection and Anti-inflammatory properties of characterized melanin produced by marine proteobacterium Providencia rettgeri strain BTKKS1 was explored in the study. Characterization of melanin was carried out by chemical, FTIR, proton NMR and EPR analysis. The radical scavenging property was estimated using DPPH assay and Fe 2+ chelating potential was also evaluated. Effect of melanin on the activities of Cyclooxygenase, Lipoxygenase, Myeloperoxidase and Cellular Nitrite is used to evaluate anti-inflammatory potential. Enhancement of Sun Protection Factor (SPF) is evaluated to study its effectiveness in photoprotection. Cytotoxicity of melanin was estimated using MTT assay.The chemical, FTIR, proton NMR and EPR characterization were typical of eumelanin. The pigment also showed profound radical scavenging activity (63.73%) and metal chelating potential (97.09%). Melanin significantly inhibited the activity of the inflammatory enzymes in a dose dependent manner and enhanced the SPF value of commercial sunscreens at an average of 2.64 factors. This melanin was also less cytotoxic with an IC50 value of 97.87μg/mL. The immense Anti-inflammatory property of the pigment can be utilized in therapeutic applications. The photoprotection potential of melanin can be utilized in cosmetic formulations, UV protection devices etc.
KEYWORDS: Anti-Inflammatory;bioactivity Melanins; Providencia Rettgeri; SPF;
Download this article as:| Copy the following to cite this article: Kurian N. K, Bhat S. G. Photoprotection and Anti-Inflammatory Properties of Non–Cytotoxic Melanin from Marine Isolate Providencia Rettgeri Strain BTKKS1. Biosci Biotech Res Asia 2017;14(4). |
| Copy the following to cite this URL: Kurian N. K, Bhat S. G. Photoprotection and Anti-Inflammatory Properties of Non–Cytotoxic Melanin from Marine Isolate Providencia Rettgeri Strain BTKKS1. Biosci Biotech Res Asia 2017;14(4). Available from: https://www.biotech-asia.org/?p=28571 |
Introduction
The dark colored biopolymer complex melanin is widely distributed in nature, in all living forms, having diverse biological functions including photo protection, thermoregulation, as free radical sinks, cation chelators and antibiotics. In plants it is incorporated as strengtheners in the cell walls (Riley 1997), whereas it not only determines the skin color in humans, but also plays a significant role in protecting skin against UV damage (Huang and Chang 2012). In microorganisms, they protect against environmental stresses, with instances of increased resistances toward antibacterials in melanin producers (Lin et al. 2005), besides being involved in fungal pathogenesis (Butler and Day 1998). According to Nicolaus (1968), melanins can be sub grouped into three namely eumelanin, a brown to black pigment derived by the oxidative polymerization of precursors like tyrosine, dihydroxyphenylalanine (DOPA), dopamine and tyramine; pheomelanin, a cysteine containing yellow to red pigment with a biosynthetic pathway similar to eumelanin and the heterogeneous allomelanins, which are formed by the polymerization of di- or tetrahydrofolate via pentaketide pathway.
The common commercial application of melanin is in cosmetics such as sunscreen lotions where it acts as a photo protective component due to its UV-protective and free radicals scavenging properties (Riley 1997). Melanins act as UV-protective agents in bioinsecticide preparation like the Bacillus thuringenesis (Bt) insecticidal crystals (Wan et al. 2007; Zhang et al. 2007). The melanin producing organism can also be used in bioremediation of radioactive waste such as uranium (Turick et al. 2008) and so on. Due to the diverse application possibilities not restricted to any particular field, the study of melanins is the demand of the hour.
Numerous bacteria like Vibrio cholerae, Shewanella colwelliana (Kotob et al. 1995) and Alteromonas nigrifaciens (Ivanova et al. 1996) produce melanins, including pyomelanin producers like Pseudomonas aeruginosa (Eiko and Ohyama 1972), Shewanella. colwelliana, Vibrio cholerae, Hyphomonas sp. (Ruzafa et al. 1995; Kotob et al. 1995) and Alcaligenes eutrophus (David et al. 1996). Marine actinomycetes including Streptomyces strains reportedly use tyrosinases in melanin synthesis. Another melanin-synthesizing microbe which produces black eumelanin from L- tyrosine is Marinomonas mediterranea (Solano and Sanchez-Amat 1999). Most of these melanin producers are terrestrial in origin, while marine bacteria remain unexplored.
Bacteria of the Morganella-Proteus-Providencia group produce a yet uncharacterized brownish melanin – like- pigment on agar containing L-form of aromatic amino acids (Müller 1985). In this work the melanin produced by marine proteobacteria Providencia rettgeri strain BTKKS1 is characterized and its various biological properties of therapeutic and cosmetological importance were explored.
Materials and Methods
Chemicals, Cell Lines and Bacterial Isolates
Synthetic melanin (Sigma Chemicals Co, St Louis, USA), L-tyrosine (Himedia chemicals, Mumbai, India) and all other chemicals used were of analytical reagent grade.
RAW 264.7 and L929 cell lines were maintained in Dulbecco’s modified eagles media (Himedia, India) supplemented with 10 % FBS (Fetal Bovine serum) (Invitrogen, USA) and grown to confluence at 37°C at 5 % CO2 in a CO2 incubator (Eppendorf, Germany).
The melanin producing Providencia rettgeri strain BTKKS1 was isolated from marine sediments from Kanyakumari (8° 5’N, 77° 32’E) coast of southern India. Screening for melanin production was initially by a plate based assay (Kurian et al. 2014) and then in tyrosine basal broth (Eiko and Ohyama 1972). The bacterium was identified by biochemical and 16SrDNA sequencing (Mac Faddin 1976; Ausbel et al. 1995; Sambrook et al. 1989; Shivaji et al. 2000).
Production, Extraction and Purification of Melanin
Tyrosine basal broth (Eiko and Ohyama 1972) containing 0.2% tyrosine was used for melanin production.5 mL of this culture suspension (OD600 = 1) was used as primary inoculums for 50 mL of production medium and kept in an environment shaker (Orbitek, Scigenics, India) at 140 rpm at 37±2oC for180 h. Melanin production kinetics was studied by sampling at 12 h intervals and estimating bacterial growth and melanin production spectrophotometrically (Turick et al. 2002).
After 180 h of incubation, the cell free supernatant was acidified to pH 2 using 1 N HCl. Black precipitate of melanin can be visualized at the bottom of the flask at lower pH . Further treatment with acid, water and ethanol simultaneously according to Sajjan et al (2013) helped to get pure melanin.
Physicochemical Characterization of Melanin
Reactivity of melanin with various organic solvents, acidic and basic solutions, oxidising and reducing agents were evaluated (Fava et al. 1993). Spectroscopic techniques such as FT-IR (Ravishankar et al. 1995), Proton NMR (Guo et al. 2014) and EPR spectroscopy (Enochs et al. 1993) were used to evaluate the biophysical properties of the pigment, as also elemental analysis (Sajjan et al. 2013). Antioxidant and metal chelating properties of the pigment was evaluated using standard procedures (Liyana-Pathirana and Shahidi 2005; Dinis et al. 1994).
Anti- Inflammatory Potential of Melanin
RAW 264.7 cells were then grown to 60% confluence followed by activation with 1µL Lipopolysaccharide (LPS) (1µg/mL). LPS stimulated RAW cells were exposed to different concentration (6.25, 12.5, 25, 50, 100 µg/mL) of melanin solution. Diclofenac sodium, a standard anti-inflammatory drug, in varying concentration corresponding to the sample was also added and incubated for 24 hours. After incubation the anti-inflammatory assays were performed using the cell lysate. Activities of three inflammatory enzymes namely Cyclooxygenase (COX) (Walker and Gierse. 2010), Lipoxygenase (LOX) (Axelrod et al. 1981), Myeloperoxidase (MPO) (Bradley et al. 1982) and Cellular nitrite levels (Lepoivre et al. 1990) were assayed using standard protocols.
Photo Protective Nature of Melanin
Photoprotective nature of melanin was expressed by its ability to enhance the Sun Protection Factor (SPF) of commercial sun screens.Sun Protection Factor (SPF) was estimated by a modified protocol (Suryawanshi et al. 2015). Commercial sunscreens of (0.1 g) was added each to 10 mL of absolute ethanol, as also melanin at 0.005% concentration. Absorbance of the mixture in the UV range (290–320 nm) was taken at 5 nm intervals using ethanol as the blank.
SPFs were calculated, according to Mansur et al. 1986, using following formula
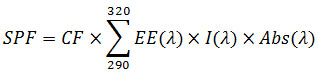
Where CF (correction factor) = 10; EE (λ) = erythmogenic effect of radiation with wavelength k; Abs (λ) = spectrophotometric absorbance value of the solution; and I = solar intensity spectrum. EE (λ) ×I is constant and was determined (Sayre et al. 1979).
Cytotoxicity of Melanin
Different concentrations (6.25, 12.5, 25, 50 and 100 μg/mL) of melanin were added to L929 cells at and incubated for 24 hours. The percentage difference in viability was determined by standard 3-(4, 5dimethythiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay (Arung et al. 2009) after 24 hours of incubation.
Statistical Analysis
All the experiments were repeated thrice. The statistical analysis was done by ANOVA using GraphPad Prism. Ver.6 computer program, where p values<0.05 were considered significant.
Results
Strain Identification
Following preliminary screening, bacteria from marine sediment sample producing a clearing zone on tyrosine agar plates were selected as melanin producers. Strain BTKKS1 selected for further characterization after secondary screening was identified as Gram negative rod, indole, methyl red and citrate positive and Voges– Proskauer negative. The bacterium was catalase positive but oxidase negative and could utilize sugars such as glucose, adonitol and manitol in the medium. It was identified further as Providencia rettgeri (KF515633) by 16S rDNA sequence analysis.
Pigment Production
Strain BTKKS1 produced considerable amount of pigment in the tyrosine broth from third day until the eight day, when pigment concentration was 30.31±0.69 μg/mL, with no further increase in production thereafter (Fig. 1).
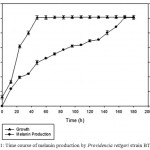 |
Figure 1: Time course of melanin production by Providencia rettgeri strain BTKKS1
|
Physicochemical Characterization of Melanin
Melanin from strain BTKKS1 was soluble in alkaline solvents like sodium hydroxide, potassium hydroxide and Dimethyl sulfoxide (DMSO). However, the pigment showed least solubility in water and common organic solvents. Oxidizing (30% H2O2) and reducing (Na2SO3) agents decolorized the pigment.
The IR spectrum showed characteristic peaks (Laxmi et al. 2016) showing similarity with those in earlier reports (Selvakumar et al. 2008). 1H NMR peaks of melanins (Fig.2) showed similarity with earlier reports (Arun et al. 2015; Guo et al. 2014) with signals in both the aromatic (7.03-7.32 ppm) and aliphatic regions (0.8 ppm). Sharp peaks in the EPR spectra (Fig. 3) of melanins indicated the presence of unpaired electrons, which can trap free radicals. This was further confirmed by the immense radical scavenging activity (63.73%) and metal chelating potential (97.09%) of the pigment at its higher concentration (100μg/mL) tested (Fig. 4).
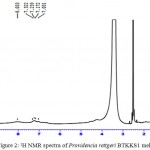 |
Figure 2: 1H NMR spectra of Providencia rettgeri BTKKS1 melanin
|
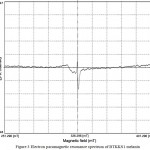 |
Figure 3 Electron paramagnetic resonance spectrum of BTKKS1 melanin
|
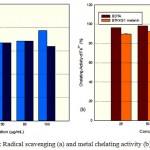 |
Figure 4: Radical scavenging (a) and metal chelating activity (b) of melanin
|
Elemental composition of Providencia rettgeri strain BTKKS1 melanin showed 47.48% carbon, 4.10% hydrogen, 12.73% nitrogen and 0.89% sulfur. Typical elemental composition of this bacterial melanin was similar to those obtained in earlier reports (Hong and Simon 2006).
Anti- Inflammatory Potential of Melanin
P. rettgeri melanin significantly inhibited the activity of four inflammatory enzymes assayed in the study in a dose dependent manner. Melanin inhibited COX at an IC50 of 95.09%, with maximum inhibition at highest concentration tested (100 μg/mL) being 52.58% (Fig. 5 a), while it showed 63.62% inhibition (IC50= 78.59 μg/mL) of LOX enzyme (Fig. 5 b).About 74 % of the MPO activity was inhibited by BTKKS1 melanin at 100 μg/mL concentration (Fig.5 c), while the cellular nitrite level also decreased considerably. (Fig. 5 d),
 |
Figure 5: Effect of BTKKS1 melanin on inflammatory enzymes (a) Cyclooxygenase (COX) (b) Lipoxygenase (LOX) (c) Myeloperoxidase (MPO) (d) Cellular Nitrite Levels
|
Photo Protective Nature of Melanin
SPF value of the sun screens tested was increased by the addition of 0.005% melanin. BTKKS1 melanin enhanced the SPF value by an average of 2.64 factors (Table 1).
Table 1: Sun protection factors (SPFs) for commercial sunscreen preparations before and after supplementation with BTKKS1 melanin
| Commercial Sunscreen | SPF value stated by the manufacturer | SPF value determined empirically during the current study | +BTKKS1 Melanin (0.005% w/w) SPF |
| Sunscreen 1 | 15 | 14.24±0.007 | 17.09±0.06 |
| Sunscreen 2 | 15 | 14.61±0.01 | 17.74±0.02 |
| Sunscreen 3 | 15 | 14.77±0.05 | 17.57±0.05 |
| Sunscreen 4 | 17 | 16.47±0.04 | 19.52±0.01 |
| Sunscreen 5 | 30 | 26.26±0.04 | 28.90±0.05 |
Cytotoxicity of Melanin
Providencia rettgeri BTKKS1 (Fig. 6) melanin was observed to be less toxic to L929 cells with an IC50 value of 97.87 μg/mL.
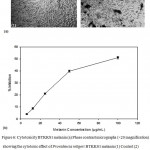 |
Figure 6: Cytotoxicity BTKKS1 melanin (a) Phase contrast micrographs (×20 magnification) showing the cytotoxic effect of Providencia rettgeri BTKKS1 melanin (1) Control (2) melanin treated (100μg/mL) (b) Graph showing the percentage inhibition of growth of L929 cells.
|
Discussion
Melanin production of BTKKS1 started from the second day and continued till day eight, when it stabilized. Earlier reports showed that melanin production in bacteria usually starts in 24-72 hours after inoculation (Zhang et al. 2007). The actual period may vary with the genus, but we don’t have many reports on melanin production by other Providencia rettgeri so to compare with BTKKS1 production pattern.
Chemical nature of melanin, especially its insolubility in most of solvents including water may be due to aromatic rings and carboxylic acids, which could get fully protonated when contacted with water. But it is solubilized in alkaline solvents and DMSO. Solubility in DMSO may be the result of thioalkylation of the phenolic units in melanins (Hansen et al. 2011).
One of the most unusual features of melanin is its persistent EPR signal (Blois et al. 1964). Indeed, melanin was among the first biological materials examined by EPR spectroscopy (Commoner et al. 1954). Melanin free radicals are stable, and the content of melanin free radicals and their corresponding EPR signal intensity can easily be modified by a number of physicochemical agents (Sealy et al. 1980) like metal ions, light etc. Ability of melanin to interact with stable free radical DPPH indicated further the scavenging activity of the pigment due to the presence of paramagnetic centres (PMC). BTKKS1 melanin was proved to bind tightly to reactive metals like Fe(II) which enables protection from Femton reactions which cause tissue damages (Flora 2009). This protective nature can be utilized in many useful applications.
Classification of melanin as pheomelanin subclass can be done by CHN(S) elemental analysis. Pheomelanin (Ito and Fujita 1985) with cysteine incorporated structure have more sulfur content (9.78%) compared to other types like synthetic dopa melanin (Ito and Fujita 1985) (0.09%) and Klebsiella sp melanin (Sajjan et al. 2013) (0.86%). The low sulfur (0.89%) content of BTKKS1 pigment contraindicated pheomelanin class (Sajjan et al. 2013).
BTKKS1 melanin decreased the activity of all inflammatory enzymes (COX, LOX, MPO and NO synthase) tested. Kurian et al. (2015) reported similar effect of Bacillus melanin. There are no other reports so far though there are many reports available regarding phenolic compounds (Masuda et al. 2010; Kato et al. 2003; Tsao et al. 2005) inhibiting the activity of these inflammatory enzymes. May be similar mechanism is also employed here. There are only few reports regarding the anti-inflammatory properties of melanin. Avramidis et al. (1998) reported that grape melanin interfered with the prostaglandin as well as the leukotriene and/or complement system mediated inflammation.
Immense improvement of photoprotection by BTKKS1 melanin supplemented sunscreens, opens doors for more melanin based cosmetics. Huang et al. (2011) reported the sun protection effect of melanin from berry of Cinnamomum burmannii and Osmanthus fragrans. Later Tarangini and Mishra (2014) also reported the profound enhancement in SPF value by Bacillus safensis melanin. The less cytotoxic nature of BTKKS1 melanin also makes it a suitable candidate for cosmetic formulations.
Thus the characterized melanins from Providencia rettgeri strain BTKKS1 had shown immense bioactivities which can be utilized further in different areas of life activities. Its anti-inflammatory properties can be utilized for therapeutic applications. While its property of SPF enhancement in sun screens makes it an essential ingredient in cosmetic formulations. More in vivo and clinical trials were required to confirm its utility
Acknowledgements
First author acknowledges DST (Dept of Science and Technology, Govt. of India) for the DST INSPIRE- Junior and Senior Research Fellowship
Conflict of Interest
Authors declare no conflict of interests
Funding Source
There is no funding sources
References
- Riley P.A. Melanin. Int J Biochem Cell Biol. 1997;29(11):1235-1239.
CrossRef - Huang H.C, Chang T.M. Antioxidative properties and inhibitory effect of Bifidobacterium adolescentis on melanogenesis. World J Microbiol Biotechnol. 2012;28(9):2903-2912.
CrossRef - Lin W.P, Lai H.L, Liu Y.L, Chiung Y.M, Shiau C.Y, Han J.M. Effect of melanin produced by a recombinant Escherichia coli on antibacterial activity of antibiotics. J Microbiol Immunol Infect. 2005;38(5):320.
- Butler M.J, Day A.W. Fungal melanins: a review. Can J Microbiol. 1998;44(12):1115-1136.
CrossRef - Nicolaus R.A. Melanins: Hermann: Paris. 1968.
- Wan X, Liu H.M, Liao Y, Su Y, Geng J, Yang M.Y, Shen P. Isolation of a novel strain of Aeromonas media producing high levels of DOPA‐melanin and assessment of the photoprotective role of the melanin in bioinsecticide applications. J Appl Microbiol. 2007;103(6):2533-2541.
CrossRef - Zhang J, Cai J, Deng Y, Chen Y, Ren G. Characterization of melanin produced by a wild-type strain of Bacillus cereus. Front Biol. 2007;2(1):26-29.
CrossRef - Turick C, Knox A, Leverette C, Kritzas G. In situ uranium stabilization by microbial metabolites. EnvironRadioact. 2008;99:890-899.
CrossRef - Kotob S, Coon S.I, Quintero E.J, Weiner R.M. Homogentisic acid is the primary precursor of melanin synthesis in Vibrio cholerae, a Hyphomonas strain and Shewanella colwelliana. Appl Environ Microbiol. 1995;61:1620-1621.
- Ivanova E.P, Kiprianova E.A, Mikhailov V.V, Levanova G.F, Garagulya A.D, Gorshkova N.M, Yumoto N, Yoshikawa S. Characterization and identification of marine Alteromonas nigrifaciens strains and emendation of the description. Int J Syst Bacteriol. 1996;46:223–228.
CrossRef - Eiko Y, Ohyama A. Characterization of “Pyome1anin”-Producing strains of Pseudomonas aeruginosa. J of Syst Bacteriol. 1972;2:53-64.
- Ruzafa C, Sanchez-Amat A, Solano F. Characterization of the melanogenic system in Vibrio cholerae, ATCC 14035. Pigment Cell Res. 1995;8:147–152.
CrossRef - David C, Daro A, Szalai E, Atarhouch T, Mergeay M. Formation of polymeric pigments in the presence of bacteria and comparison with chemical oxidative coupling; II. Catabolism of tyrosine and hydroxyphenylacetic acid by Alcaligenes eutrophus CH34 and mutants. Eur Polym J. 1996;32:669–679.
CrossRef - Solano F, Sanchez-Amat A. Studies on the phylogenetic relationships of melanogenic marine bacteria: proposal of Marinomonas mediterranea Nov. Int J Syst Bacteriol. 1999;49:1241-1246.
CrossRef - Müller H.E. Production of brownish pigment by bacteria of the. Morganella-Proteus-Providencia Zentralbl Bakteriol Mikrobiol Hyg A. 1985;260(4):428-35.
CrossRef - Kurian N.K, Nair H.P, Bhat S.G. Melanin producing Pseudomonas stutzeri BTCZ10 from marine sediment at 96 m depth (Sagar Sampada cruise #305). Int J.Curr.Biotechnol. 2014;2(5):6-11.
- MacFaddin J.F. Biochemical tests for identification of medical bacteria. Williams & Wilkins Co. 1976.
- Ausbel F.M, Brent R, Kingston R.E, Moore D.D, Seidman J.G, Smith J.A, Struhl K. Short protocols in molecular biology. New York: John Wiley & Sons. 1995.
- Sambrook J, Fritsch E.F, Maniatis T. Molecular cloning .New York: Cold spring harbor laboratory press. 1989.
- Shivaji S, Bhanu N.V, Aggarwal R.K. Identification of Yersinia pestis as the causative organism of plague in India as determined by 16S rDNA sequencing and RAPD-based genomic fingerprinting. FEMS Microbiol Lett. 2000;189(2):247-252.
CrossRef - Turick C.E, Tisa L.S, Caccavo J.F. Melanin production and use as a soluble electron shuttle for Fe(III) oxide reduction and as a terminal electron acceptor by Schewanella algae Appl. Environ. Microbiol. 2002;68:2436-2444.
CrossRef - Sajjan S.S, Anjaneya O, Kulkarni G.B, Nayak A.S, Mashetty S.B, Karegoudar T.B. Properties and Functions of Melanin Pigment from Klebsiella GSK. Korean J. Microbiol. Biotechnol. 2013;41(1):1-10.
CrossRef - Fava F, Gioia D.D, Merchetti L. Characterization of a pigment produced by Pseudomonas fluorescens during 3- chlorobenzoate co-metabolism. 1993;27:825-835.
- Ravishankar J.P, Muruganandam V, Suryanarayanan T.S. Isolation and characterization of melanin from a marine fungus. Botanica Marina. 1995;38:413-416.
CrossRef - Guo J, Rao Z, Yang T, Man Z, Xu M, Zhang X. High-level production of melanin by a novel isolate of Streptomyces kathirae. FEMS Microbiol Lett. 2014;357(1):85-91.
CrossRef - Enochs W.S, Nilges M.J, Swartz H.M. A standardized test for theidentification and characterization of melanins using electronparamagnetic resonance (EPR) spectroscopy. Pigment Cell 1993;6(2):91-99.
CrossRef - Liyana-Pathirana C.M., Shahidi F. Antioxidant activity of commercial soft and hard wheat (Triticum aestivum ) as affected by gastric pH conditions. J Agric Food Chem. 2005; 53(7):2433-2440.
CrossRef - Dinis T.C, Madeira V.M, Almeida L.M. Action of phenolicderivatives (acetaminophen, salicylate, and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Biochem. Biophys. 1994;315(1):161-169.
CrossRef - Walker M.C, Gierse J.K. In vitro assays for cyclooxygenase activity and inhibitor characterization. In Humana Press. 2010;131-144.
CrossRef - Axelrod B, Cheesbrough T.M, Laakso S. Lipoxygenase from soybeans: EC 1.13. 11.12 Linoleate: oxygen oxidoreductase. Methods Enzymol. 1981;71[53]:441-451.
CrossRef - Bradley P.P, Priebat D.A, Christensen R.D, Rothstein G. Measurement of cutaneous inflammation: estimation of neutrophil contentwith an enzyme marker. J. Invest. Dermatol. 1982;78(3):206-209.
CrossRef - Lepoivre M, Chenais B, Yapo A, Lemaire G, Thelander L, Tenu J.P. Alterations of ribonucleotide reductase activity following induction of the nitrite-generating pathway in adenocarcinoma cells. Biol. Chem. 1990;265(24):14143-14149.
- Suryawanshi R.K, Patil C.D, Borase H.P, Narkhede C.P, Stevenson A, Hallsworth J.E, Patil S.V. Towards an understanding of bacterial metabolites prodigiosin and violacein and their potential for use in commercial sunscreens. Int J Cosmet Sci. 2015;37(1):98-107.
CrossRef - Mansur J.D.S, Breder M.N.R, Mansur M.C.D.A, Azulay R.D. Determinaçäo do fator de proteçäo solar porespectro fotometria. Bras.Dermatol. 1986;61(3):121-124.
- Sayre R.M, Agin P.P, LeVee G.J, Marlowe E. A. comparison of in vivo and in vitro testing of sunscreening formulas. Photochem. Photobiol. 1979;29(3):559-566.
CrossRef - Arung E.T, Wicaksono B.D, Handoko Y.A, Kusuma I.W, Yulia D, Sandra, F. Anti-cancer properties of diethylether extract of wood from sukun (Artocarpus altilis) in human breast cancer (T47D)cells. Trop J Pharm Res. 2009;8(4).
- Laxmi M, Kurian N.K, Smitha S, Bhat S.G. Melanin and bacteriocin from marine bacteria inhibit biofilms of foodborne pathogens. Indian J. Biotechnol. 2016;15(3):392-399.
- Selvakumar, Rajasekar S, Periasamy K, Raaman N. Isolation and characterization of melanin pigment from Pleurotus cystidios (telomorph of Antromyocopsis macrocapa). World J. Microbiol. Biotechnol. 2008;24:2125-2131.
CrossRef - Arun G, Eyini M, Gunasekaran P. Characterization and biological activities of extracellular melanin produced by Schizophyllum commune (Fries). Indian J Exp Biol. 2015;53(6):380-387.
- Hong L, Simon J.D. Insight into the binding of divalent cations to Sepia eumelanin from IR absorption spectroscopy. Photochem Photobiol. 2006;82:1265-1269.
CrossRef - Hansen S, Pedersen-Bjergaard S, Rasmussen K. Introduction to pharmaceutical chemical analysis. John Wiley & Sons. 2011;302-310.
CrossRef - Blois M.S, Zahlan A.B, Maling J.E. Electron spin resonance studies on melanin. J. 1964;4(6):471-490.
CrossRef - Commoner B, Townsend J, Pake G.E. Free radicals in biological materials. 1954;174(4432):689.
- Sealy R.C, Felix C.C, Hyde J.S, Swartz H.M. Structure and reactivity of melanins: influence of free radicals and metal ions. Free radicals in biology. 1980;4:209-259.
CrossRef - Flora S.J. Structural, chemical and biological aspects of antioxidants for strategies against metal and metalloid exposure. Oxid Med Cell Longev. 2009;2(4):191-206.
CrossRef - Ito S, Fujita K. Microanalysis of eumelanin and pheomelanin in hair and melanomas by chemical degradation and liquid chromatography. Biochem. 1985;144:527-536.
CrossRef - Kurian N.K, Nair H.P, Bhat S.G. Evaluation of Anti-inflammatory property of Melanin from marine Bacillus sp BTCZ31. Asian J Pharm Clin Res. 2015;8(3):251-255.
- Masuda T, Someya T, Fujimoto A. Phenolic inhibitors of chemical and enzymatic oxidation in the leaves of Myricarubra. Biotechnol. Biochem. 2010;74(1):212-215.
CrossRef - Kato Y, Nagao A, Terao J, Osawa T. Inhibition ofmyeloperoxidase-catalyzed tyrosylation by phenolic antioxidants in vitro. Biotechnol. Biochem. 2003;67(5):1136-1139.
CrossRef - Tsao L.T, Tsai P.S, Lin R.H, Huang L.J, Kuo S.C, Wang J.P. Inhibition of lipopolysaccharide-induced expression of inducible nitric oxide synthase by phenolic (3E)-4-(2-hydroxyphenyl) but-3-en-2-one in RAW 264.7 macrophages. Biochem Pharmacol. 2005;70(4):618-626.
CrossRef - Avramidis N, Kourounakis A, Hadjipetrou L, Senchuk V. Antiinflammatory and immunomodulating properties of grape melanin. Inhibitory effects on paw edema and adjuvant induced disease. Arzneimittel-Forschung. 1998;48(7):764-771.
- Huang S, Pan Y, Gan D, Ouyang X, Tang S, Ekunwe S.I, Wang H. Antioxidant activities and UV-protective properties of melanin from the berry of Cinnamomum burmannii and Osmanthus fragrans. Med Chem Res. 2011;20(4):475-481.
CrossRef - Tarangini K, Mishra S. Production of melanin by soil microbial isolate on fruit waste extract: two step optimization of key parameters. Biotechnol Rep. 2014;4:139-146.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.



