

How to Cite | Publication History | PlumX Article Matrix
Charul1 , Sunali Bandral1, Shivalika1, Vikas Dogra2, Mohd. Feroz3 , Umer Bin Farook4 and Rakesh K. Panjaliya1*
, Sunali Bandral1, Shivalika1, Vikas Dogra2, Mohd. Feroz3 , Umer Bin Farook4 and Rakesh K. Panjaliya1*
1Department of Zoology, University of Jammu, J and K, India.
2Govt. College for Women, Parade, Jammu, J and K, India.
3Department of Zoology, AAAM Degree College Bamina, Srinagar, J and K, India.
4Faculty of Agriculture, SKAUST, Kashmir, India.
Corresponding Author E-mail: rakeshpanjaliya@jammuuniversity.ac.in
DOI : http://dx.doi.org/10.13005/bbra/3073

ABSTRACT: Hymenoptera is the fourth diverse and the most economically important insect order comprising of bees, wasps, ants, sawflies etc. Being an important part of ecosystem, their conservation is of utmost importance. The first step towards conservation strategies is the identification of the species. The traditional morphological approach can sometimes lead to misidentification due to a lack of expertise. DNA barcoding using the small genomic fragments has been identified as an efficient tool in the identification as well as the phylogenetic analysis of the species. In the present study, we used the COI gene sequences as a tool for the characterization of Hymenoptera from different parts of the Jammu region. The collected samples were proceeded for the isolation of DNA, PCR for amplification of the COI gene, and then sequenced by Sanger dideoxy method. A total of 22 COI sequences belonging to 18 different species were successfully generated. Among which eight species sequences (Tachytes sp., Bombus trifasciatus, Rhynchium carnaticum, Ropalidia brevita, Lasioglossum marginatum, Camponotus pennsylvanicus, Tapinoma melanocephalum, Formica rufibarbis) are the novel contribution in the global database. NJ tree using the K2P model with 1000 bootstrap supporting values has been used to study the phylogeny of the species. Sequence analysis shows high AT content (67-77%) in the COI region of Hymenopterans. The generated COI sequence analysis also revealed less than 1% intra-specific divergence in the examined taxa, while the interspecific distances ranged between 8% to 38%. This study added significantly to the databases of DNA barcodes of Hymenopterans species from Jammu region.
KEYWORDS: COI gene; DNA barcoding; Hymenoptera; Phylogenetic study
Download this article as:| Copy the following to cite this article: Charul C, Bandral S, Shivalika S, Dogra V, Feroz M, Farook U. B, Panjaliya R. K. Phylogenetic Analysis of the Economically Important Hymenopterans using cytochrome oxidase 1 Enzyme Sequences. Biosci Biotech Res Asia 2023;20(1). |
| Copy the following to cite this URL: Charul C, Bandral S, Shivalika S, Dogra V, Feroz M, Farook U. B, Panjaliya R. K. Phylogenetic Analysis of the Economically Important Hymenopterans using cytochrome oxidase 1 Enzyme Sequences. Biosci Biotech Res Asia 2023;20(1). Available from: https://bit.ly/3WPJcgt |
Introduction
Hymenoptera is one of the four mega-diverse insect orders after Coleoptera, Lepidoptera, and Diptera, comprising up to a million undescribed extant species and 153,000 described ones1, 2. The order includes ants, bees, wasps, etc. which are of great worth to all terrestrial ecosystems and of economic importance for humanity3, 4. Bees and some species of wasps are the most effective pollinators of agricultural and wild plants and help sustain our food supplies4, 5, 6. Various wasp species are the predators or parasitoids of crop pests and can be used as a substitute for chemical pesticides have benefits for the environment and are cost-effective4, 7, 8, 9. The importance of ants in the ecosystem is also widespread due to their interactions with different plants, feeding on other insects and small invertebrates, seed dispersal, and seed-predator habit10, 11.
A sharp decline in the population of these insects has been observed globally due to anthropogenic activities like deforestation, urbanisation, and application of pesticides in agricultural fields, leading to various adverse effects like habitat fragmentation, climate change and degradation of the insect’s habitat10, 11. For the maintenance of biodiversity and ecological integrity and considering the spanning human needs and interests of these creatures, their conservation is the need of the hour. And the first step in conservation is the accurate identification of the species. The traditional taxonomic methods used for the identification require the expertise of the field with a thorough knowledge of taxonomic terms to avoid misidentifications. Not only this, the technique has limitations in the identification of morphologically cryptic species.
Moreover, the available keys also work for a particular life stage or sex12. To overcome the limitations of the morphological taxonomic approach, a molecular technique involving the generation of species-specific barcodes was introduced. Hebert et al., 2003 first established the use of the Mitochondrial COI gene to generate bio-identification barcodes in eukaryotic animals. The mitochondrial COI gene is considered an effective ‘barcode’ in species identification and delimitation because of its faster rate of evolution, which helps in the study of closely related taxa that have recently diverged13, 14, 15. The present study aims to generate the DNA barcodes for the Hymenoptera species from the Jammu region to study phylogenetic relations among them. The study helps in estimating hymenopteran diversity in the region and helps formulate conservation strategies for this species diversity.
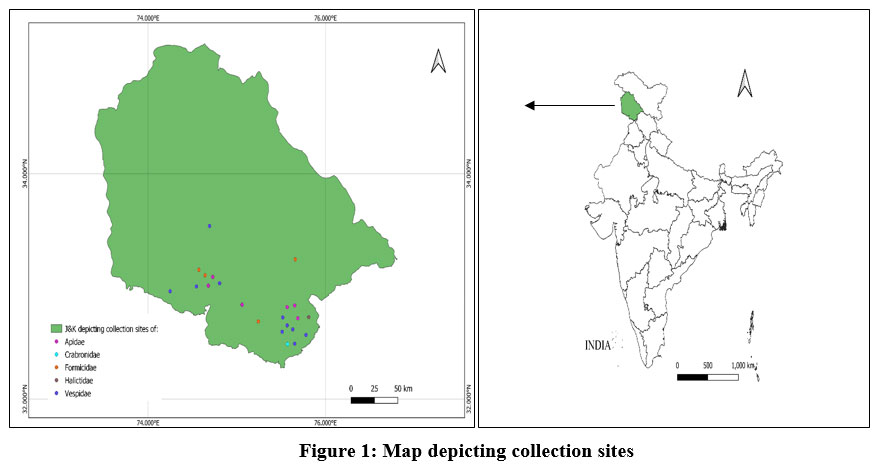 |
Figure 1: Map depicting collection sites |
Materials and methods
Taxon sampling and vouchering
About 53 samples were collected from different areas of the Jammu region by using sweeping nets and handpicking. The insects were anesthetised using ethyl acetate and preserved in 70% molecular grade alcohol for both morphological and molecular examination. The samples were examined under the Olympus SZ2-ILST for morphological identification, and voucher IDs were assigned to them.
Molecular interrogations, sequence annotation and dataset preparation
For DNA isolation, 1-2 legs depending upon the size and finally chopped, are used in the case of bees and wasps, while an intersegmental abdominal cut was made in the case of ants. The tissue was lysed overnight in buffer ATL and proteinase-K at 56ºC with 150rpm (revolutions per minute) in Stvart SBS40 shaking water bath (Cole- Paramer Ltd. Stone, ST15 OSA, UK). After tissue lysis, the total genomic DNA was extracted by using DNeasy Blood & Tissue Kit (Hilden, Germany). Polymerase Chain Reaction (PCR) was performed in applied biosystems ProFlex PCR system by life technologies to amplify partial mitochondrial cytochrome oxidase subunit I (COI) gene by using primers and conditions as shown in Table 1. The PCR products were sent to Biologia Research India Pvt. Ltd. for the sanger sequencing. The generated forward and reverse chromatogram files for each specimen were checked in MEGA X16 . The generated sequences were further screened in BLASTn (Basic Local Alignment Search Tool) for the available similar sequences in NCBI (National Centre for Biological Information) and 10 similar database sequences of each species are obtained from NCBI and are used for data curation. ClustalW software17 has been used for pairwise and multiple alignments by using a Gap opening penalty of 15.00 and a Gap extension penalty of 6.66 to obtain the final sequence for submission. The final dataset of 22 sequences from 18 different species was submitted to the NCBI Genbank through BioEdit; an online submission portal and the GenBank accession numbers were generated for them, written in Table 2. The A, T, G, C, AT, and GC content of all the sequences were obtained by using science buddies, and online software and their percentages present in sequence are depicted in Table 3.
Table 1: Primer sequences used
|
Gene Fragment |
Fragment size |
Direction |
Published name |
Sequence 5’- 3’ |
References13, 50 |
Used concentration of PCR components |
Reaction conditions |
|
COI |
650bp |
Forward |
LCO 1490 |
5’-GGTCAACA |
Folmer et al., 1994 |
1x Dream Taq buffer, 2.5mM MgCl2, 0.2mMdNTP mix, 0.2μM each forward and reverse primer, 1.5U Dream Taq DNA polymerase. |
Initial denaturation at 94˚C for 4 minutes. Denaturation at 94˚C for 30 s. Annealing at 47˚C for 45 s. Extension at 72˚C for 45 s. No. of cycles= 35. Final extension at 72˚C for 20 minutes. |
|
Reverse |
HCO 2198 |
5’-TAAACTTCAG |
|
||||
|
Forward |
Lep-F1 |
5’-ATTCAACCAAT |
Hebert et al., 2004a |
0,9x Buffer, 2mM MgCl2, 0.2mM dNTP mix, 0.2mM each forward and reverse primer, 1.5U Taq polymerase |
Initial denaturation at 94˚C for 4 minutes. Denaturation at 94˚C for 30 s. Annealing at 47˚C for 45 s. Extension at 72˚C for 45 s. No. of cycles= 35. Final extension at 72˚C for 20 minutes. |
||
|
Reverse |
Lep-R1 |
5’-TAAACTTCTG |
Genetic divergence and phylogenetic tree interpretation
The pairwise genetic distance was analysed using the Kimura 2-Parameter (K2P) model in MEGA X16, 18. The number of base substitutions per site was analysed between all 22 sequences. Codon positions included were 1st+ 2nd+ 3rd+non-coding. All ambiguous positions were removed for each sequence pair, and a total of 538 positions were present in the final dataset. The evolutionary relationship was inferred using the Neighbor-Joining (NJ) method and Kimura 2- parameter model with 1000 bootstrap replications in MEGAX16, 18, 19, 20. The database sequence of Eristalis tenax (ON210045) under order Diptera was used as an out-group in the phylogenetic study.
Results and discussion
Morphological identification
The hymenopteran species were identified by consulting the available keys in the literature by Archer (2014), Bingham (1897), Carpenter and Thi Nguyen (2003), Das and Gupta (1989), Goulet and Huber (1993), Kumar and Carpenter (2013), Kumar and Srinivasan (2010), Niup and Dorji (2016), Saini et al. (2011), Williams (1998), other relevant literature and identification keys21-30. After morphological identification, the collected specimens were identified as18 different species belonging to 5 families, namely Apidae, Formicidae, Vespidae, Halictidae, and Crabronid. The details of identified species, their collection date, the site of collection & their coordinates have been listed in Table 2.
Table 2: Information about studied samples
|
Order |
Family |
Identified species name |
Voucher specimen |
Date of collection |
Place of collection |
Latitude |
Longitude |
Elevation (metres) |
Genbenk Accession number generated (COI) |
|
Hymenoptera |
Apidae |
Apis cerana |
RHY10 |
29/08/2019 |
Hostel,University of Jammu |
32.7180N |
74.8702E |
319 |
ON259474 |
|
Apis dorsata |
RHY12 |
29/08/2019 |
Hostel, University of Jammu |
32.7180N |
74.8702E |
320 |
ON259323 |
||
|
Apis dorsata |
RHY51 |
26/12/2020 |
Kathua |
32.7329N |
74.8642E |
352 |
ON259531 |
||
|
Apis mellifera |
CH9 |
6/11/2021 |
Vijaypur |
32.7053N |
74.8800E |
341 |
ON259532 |
||
|
Bombus trifasciatus |
RHY7 |
25/08/2019 |
Ramkot |
32.6042N |
75.2881E |
516 |
ON306321 |
||
|
Bombus trifasciatus |
RB9A |
23/06/2019 |
Ramkot |
32.6101N |
75.2822E |
505 |
ON306319 |
||
|
Formicidae |
Camponotus pennslyvanicus |
ANT 0 |
27/12/2021 |
Department of Zoology, University of Jammu. |
32.7242N |
74.8666E |
316 |
ON259314 |
|
|
Tapinoma melanocephalum |
ANT1 |
30/12/2021 |
Hostel, University of Jammu |
32.7180N |
74.8702E |
319 |
ON259315 |
||
|
Formica rufibarbis |
ANT3 |
10/10/2021 |
Doda |
33.1493N |
75.5477E |
1200 |
ON259316 |
||
|
Monomorium indicum |
CH6 |
29/09/2021 |
Ghaghwal |
32.5092N |
75.1863E |
366 |
ON254654 |
||
|
Vespidae |
Ropalidia brevita |
RHY2 |
25/08/2019 |
Ramkot |
32.6042N |
75.2881E |
511 |
ON306323 |
|
|
Polistes olivaceus |
RHY4 |
25/08/2019 |
Ramkot |
32.6042N |
75.2881E |
511 |
ON306320 |
||
|
Delta pyriforme pyriforme |
RHY24 |
24/10/2019 |
Ramkot |
32.6081N |
75.2786E |
453 |
ON306325 |
||
|
Vespa basalis |
RHY30 |
4/11/2019 |
Domael, Katra |
32.9395N |
74.9499E |
715 |
ON306327 |
||
|
Antodynerus limbatus |
RHY37 |
7/7/2020 |
Ramkot |
32.6103N |
75.2823E |
500 |
ON306328 |
||
|
Vespa tropica |
RHY40 |
7/7/2020 |
Ramkot |
32.6103N |
75.2823E |
500 |
ON306329 |
||
|
Polistes watti |
RHY49 |
24/12/2020 |
Campus, University of Jammu |
32.7194N |
74.8681E |
318 |
ON306330 |
||
|
Rhynchium carnaticum |
RHY50 |
20/12/2020 |
R.S. Pura |
32.7329N |
74.8642E |
351 |
ON306322 |
||
|
Vespa basalis |
RV4 |
7/4/2019 |
Hostel, University of Jammu |
32.7180N |
74.8702E |
319 |
ON306318 |
||
|
Antodynerus limbatus |
RHY22 |
24/10/2019 |
Ramkot |
32.6081N |
75.2786E |
449 |
ON306328 |
||
|
Halictidae |
Lasioglossum marginatum |
RHY 26 |
24/08/2019 |
Ramkot |
32.6081N |
75.2786E |
447 |
ON306326 |
|
|
Crabronidae |
Tachytes sp. |
RHY 1 |
25/08/2019 |
Ramkot |
32.6042N |
75.2881E |
521 |
ON306317 |
Molecular characterisati
Of the 53 collected specimens, DNA was successfully isolated from 45 samples, and successful amplification of the COI region of 35 was carried out. Of which, sequences of 22 samples have been generated, which can be accessed by the NCBI with accession numbers (ON254654; ON259314-ON259316; ON259323; ON259474; ON259531-ON259532; ON306317-ON306330). The results of the similarity search in the NCBI database showed 99-100% identical matches with the same species except for eight species (Tachytes sp., Bombus trifasciatus, Rhynchium carnaticum, Ropalidia brevita, Lasioglossum marginatum, Camponotus pennsylvanicus, Tapinoma melanocephalum, Formica rufibarbis). Hence, the present study contributed eight new sequences of hymenopteran species from the Jammu region to the database. The COI sequences were found to have less GC% (23-33%) and more AT% (67-77%), as depicted in table 3. The minimum GC% was found in Apidae and the maximum in Formicidae of the studied families of Hymenoptera. The findings are following the previous studies, which reported 22-41% GC content in the hymenopteran COI region with 22-29% in Apidae and 24-41% in formicidae31, 32. The phylogenetic tree was constructed using the NJ method and the K2P model along with 1000 bootstrap support values because the maximum literature reviewed for this research work was found to have used the NJ tree with the K2P model with supporting bootstrap values. The phylogenetic tree constructed depicts eighteen distinct lineages of twenty-two hymenopteran species belonging to five clades corresponding to the five families (Figure 2). Family Formicidae is found to be more closely related to Vespidae, while the family Apidae is found to be more closely related to the family Halictidae. The family Crabronidae is distantly related to all the four studied families; namely, apidae, formicidae, halictidae and Vespidae, as clearly depicted in the phylogenetic tree. The phylogeny constructed using COI sequences has been found as per the previous studies based on the morphological as well as molecular approach. Brothers in 1999 studied the phylogeny and evolutionary relationship of the wasps, ants and bees by using a morphological system and their results depict that ants and wasps are more closely related to each other than their relatedness to bees33. Relatedness between Formicidae and Vespidae has been reported in the studies using different molecular markers34. There is much evidence from the literature which supports the use of molecular markers for species identification and phylogenetic studies. The successful use of molecular markers especially COI for the identification and delimitation of the species was seen in Vespa species identification, Trissolcus species identification, new species of Zaischnopsis, trichogrammatids, hymenopteran parasitoids, and Egyptian wasps are some of the examples from the literature35-41. The molecular data has also been found to be useful for species delimitation as depicted by the studies of Chen and coworkers; Parslow and associates; Benavides and associates and Siddiqui and coworkers42-45. Kwon and associates in their study constructed the phylogenetic tree for the species of genus Osmia using COI gene data and found that the results were satisfactory and in accordance with the traditional approach46. Similarly, molecular characters including COI, 16S rDNA, and 28S rDNA were used to study the evolutionary transitions in parasitic wasps belonging to Apocrita in recent times47. Aman-zuki and coworkers through their work also suggested the usefulness of molecular markers in resolving the phylogeny of species belonging to Microgastrine wasps48. The studies of Niehuis and coworkers with the Euchroeus group of tribe Chrysidini have detected the inappropriate rooting of the group with the help of molecular data. Their study provides the insights that the molecular data not only supports traditional taxonomic views but also helps in resolving the phylogeny of various groups where the morphological characters have given erroneous interpretations49. Hence, the study supports the use of COI gene sequences (molecular markers) for inferring the phylogenetic relationship among the species in addition to species identification and delimitation. The genetic relatedness among the species was also calculated with the generated sequence data. The overall mean genetic distance among the species in the present dataset was found to be 26.9%. The intraspecific genetic distance ranges from 0.19% (Apis cerana) to 0.58% (Bombus trifasciatus). However, the highest inter-specific genetic distance (37.69%) was observed between Apis cerana and Monomorium indicum, and the lowest inter-specific genetic distance (8.67%) was observed between Apis cerana and Apis dorsata. The values are clearly depicted in Table
Table 3: ATGC content of all the studied Samples
|
Sample Name |
All base count |
A (%) |
T (%) |
G (%) |
C (%) |
GC (%) |
AT (%) |
|
Apis cerana |
538 |
179 |
222 |
57 |
80 |
25.5 |
74.5 |
|
Apis dorsata |
580 |
189 |
249 |
59 |
83 |
24.5 |
75.5 |
|
Apis dorsata |
538 |
178 |
227 |
57 |
76 |
24.7 |
75.3 |
|
Apis mellifera |
578 |
189 |
241 |
60 |
88 |
25.6 |
74.4 |
|
Bombus trifasciatus |
564 |
183 |
246 |
56 |
79 |
23.9 |
76.1 |
|
Bombus trifasciatus |
672 |
228 |
285 |
71 |
88 |
23.7 |
76.3 |
|
Tachytes sp. |
598 |
202 |
248 |
77 |
71 |
24.7 |
75.3 |
|
Camponotus pennsylvanicus |
637 |
191 |
268 |
71 |
107 |
27.9 |
72.1 |
|
Tapinoma melanocephalum |
654 |
194 |
246 |
86 |
128 |
32.7 |
67.3 |
|
Formica rufibarbis |
660 |
204 |
259 |
75 |
122 |
29.8 |
70.2 |
|
Monomorium indicum |
636 |
192 |
250 |
81 |
113 |
30.5 |
69.5 |
|
Lasioglossum marginatum |
621 |
197 |
251 |
70 |
103 |
27.9 |
72.1 |
|
Ropalidia brevita |
604 |
187 |
240 |
76 |
101 |
29.3 |
70.7 |
|
Polistes olivaceus |
672 |
223 |
262 |
78 |
109 |
27.8 |
72.2 |
|
Delta pyriforme pyriforme |
604 |
211 |
228 |
73 |
92 |
27.3 |
72.7 |
|
Vespa basilis |
560 |
178 |
217 |
68 |
97 |
29.5 |
70.5 |
|
Antodynerus limbatus |
604 |
185 |
246 |
79 |
94 |
28.6 |
71.4 |
|
Vespa tropica |
604 |
179 |
234 |
76 |
115 |
31.6 |
68.4 |
|
Polistes watti |
604 |
189 |
252 |
73 |
90 |
27 |
73 |
|
Rhynchium carnaticum |
598 |
172 |
244 |
78 |
104 |
30.4 |
69.6 |
|
Vespa basalis |
672 |
206 |
273 |
81 |
112 |
28.7 |
71.3 |
|
Antodynerus limbatus |
604 |
185 |
245 |
79 |
95 |
28.8 |
71.2 |
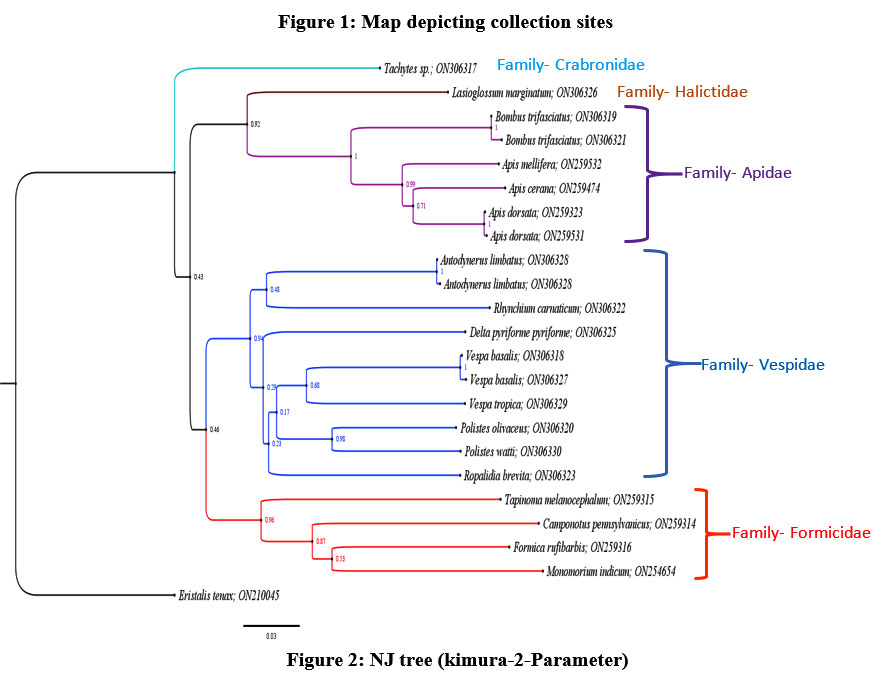 |
Figure 2: NJ tree (kimura-2-Parameter) |
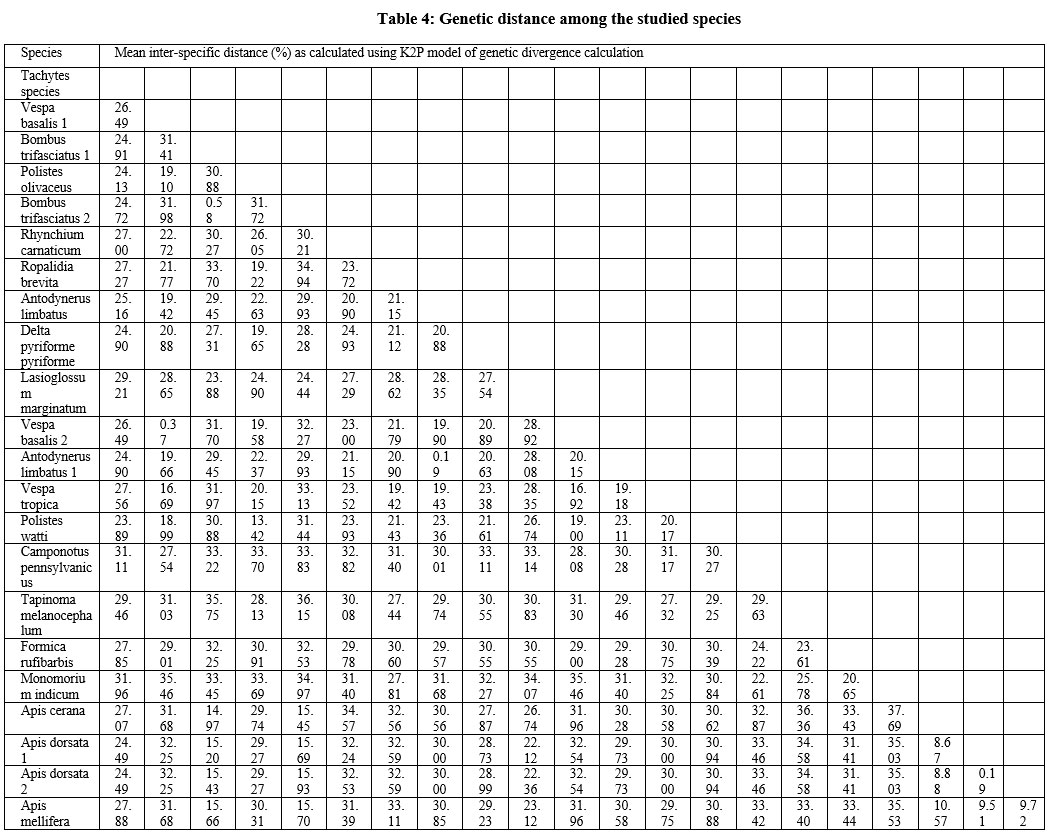 |
Table 4: Genetic distance among the studied species |
Summary and Conclusion
The present study was the first approach for carrying out molecular characterisation and phylogenetic analysis of Hymenoptera from the Jammu region. The study contributed 22 barcode sequences including 8 novel sequences belonging to the insects of order Hymenoptera to the database. The study also provided evidence in support of the use of molecular markers for phylogenetic analyses as the results are in concordance with the morphological available evidence. The current study also helps future workers, especially non-taxonomists, to identify the species based on the generated barcodes of the species to the online database.
Acknowledgement
The authors are grateful to the Head, Department of Zoology, the University of Jammu, for providing necessary facilities and equipment availability (purchased out of RUSA/ PURSE/ FIST grants). One of the authors, Charul, also acknowledges the financial support from The Department of Science and Technology, Government of India, INSPIRE Fellowship under sanction no.: DST/INSPIRE Fellowship/2018/IF180760.
Conflict of Interest
Charul carried out the extensive field work, lab work and data compilation; Sunali Bandral and Shivalika also performed the lab work; Mohd Feroz and Umer Bin Farooq identified the Hymenopteran insect species; Vikas Dogra and Rakesh K. Panjaliya has framed the work plan, provide guidance at every step and reviewed the article.
References
- Agular A.P., Deans A.R., Engel M.S., Forshage M., Huber J.T., Jennings J.T., Johnson N.F., Lelej A.S., Longino J.T., Lohrmann V., et al. Order Hymenoptera. Zootaxa. 2013; 3703:51-62.
- Grimaldi D.A., Engel M.S. Evolution of the Insects (Cambridge University Press). 2005.
- Quicke D.L.J. Parasitic Wasps (Chapman & Hall). 1997.
- Toler T.R., Evans E.W., Tepedino V. J. Pan-trapping for bees (Hymenoptera: Apiformes) in Utah’s West Desert: the importance of color diversity. Pan Pacific Entomologist 2005; 81(3-4):103-113.
- Free J.B. Insect pollination of crops. 2nd ed. Academic press.1993; 1-684.
- Raj H., Mattu V.K., Thakur M.L. Pollinator diversity and relative abundance of insect visitors on apple crop in Shimla hills of western Himalaya, India. International Journal of Science and Nature. 2012; 3(3):507-513.
- Bennett F.D., Yaseen M., Beg M.N., Sommeijer M.J. Anastrepha spp.- investigations on their natural enemies and establishment of Biosteres longicaudatus in Trinidad, West Indies. CIBC Tech. Bull. 1997; 18:1-12.
- Wong T.T.Y., Ramadan M.M., Mclnnis D.O., Mochizuki N. Influence of cohort age and host age on oviposition activity and offspring sex-ratio of Biosteres tryoni (Hymenoptera: Braconidae), a larval parasitoid of Ceratitis capitate (Diptera: Tephritidae). J. Econ. Entomol. 1990; 83: 779-783.s
- Wharton R.A. Classical biological control of fruit infesting Tephritidae. In: Robinson, A.S., Hooper, G. (Eds.), Fruit Flies: Their Biology, Natural Enemies, and Control. Elsevier, Amsterdam. 1989; pp. 303-313.
- Suryanto D.W.I. A survey of ants as candidates for potential biology control of pear psylla (Cacopsylla pyricola Foerster) in Michigan (Michigan State University) Dissertation Abstracts. 1993. MAI32/03.
- Patkar N.B., Chavan R.J. Diversity of Ants (Hymenoptera: Formicidae) from Undisturbed and disturbed habitats of Great Indian Bustard Wildlife Sanctuary, (M.S.), India. Interntional Journal of Scientific Research. 2014; 3(12):398-401.
- Hebert P.D.N., Cywinska A., Ball S.L., deWaard J.R. Biological identifications through DNA barcodes. The Royal Society of London. 2003; 270:313-321.
- Hebert P.D.N., Penton E.H., Burns J.M., Janzen D.H., Hallwachs W. Ten species in one: DNA Barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proceedings of the National Academy of Sciences of the United States of America. 2004a; 101:14812-14817.
- Ward R.D., Zemlak T.S., Innes B.H., Last P.R., Hebert P.D.N. DNA Barcoding Australia’s fish species. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 2005; 360:1847-1857.
- Smith D. Biological control of Ceroplastes rubens Maskell, by the introduced parasitoid Anicetus benefices Ishii and Yasumatsu. Queensland Journal of Agricultural and Animal Sciences. 1986; 43, 101-105.
- Kumar S., Stecher G., Li M., Knyaz C., Tamura K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Molecular Biology and Evolution. 2018; 35:1547-1549.
- Higgins D., Thompson J., Gibson T., Thompson J.D., Higgins D.G., Gibson T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994; 22:4673-4680.
- Kimura M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution. 1980; 16:111-120.
- Felsenstein J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 1985; 39:783-791.
- Saitou N., Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution. 1987; 4:406-425.
- Archer M. An alternative to the males of the Vespa species (Hymenoptera: Vespidae). Entomologist’s Monthly Magazine. 2014; 151: 57-62.
- Bingham C. T. The Fauna of British India, Including Ceylon and Burma, Hymenoptera, Vol. I, London, UK (1897).
- Carpenter J.M. and Thi Nguyen L.P. Keys to the genera of social wasps of South-East Asia (Hymnoptera: Vespiae). Entomological Science. 2003; 6: 183-192.
- Das B.P. and Gupta V.K. The social wasps of India and adjacent countries. Oriental Insects Monograph. 1989; 11:1-292.
- Goulet H., Huber J.T. Hymenoptera of the world: An identification guide to families. Centre for Land and Biological Resources Research Ottawa, Ontario. 1993; 1894-1993.
- Kumar G.P. and Carpenter J.M. A taxonomic review of the genus Antodynerus de Saussure, 1855 (Hymenoptera: Vespidae: Eumeninae) from the Indian subcontinent. Zootaxa. 2013; 3731 (2): 267-278.
- Kumar P.G. and Srinivasan G. Taxonomic studies of hornet wasps (Hymenoptera: Vespidae) Vespa Linneaus of India. Records of the Zoological Survey of India. 2010; 110(2): 57-80.
- Nidup T. and Dorji P. The Honey Bees (Hymenoptera: Apidae) of Bhutan with a key to the Apis species. Bio Bulletin. 2016; 2(2): 01-07.
- Saini M.S., Raina R.H., Khan Z.H. A Check list of Bumblebees (Hymenoptera: Apidae) from Indian Himalaya. Journal of Insect Science. 2011; 24:326-52.
- Williams P. H. Bull. Nat. History Mus. London UK. Entomol. 1998; 67: 79.
- Kim D.W., Yoo W.G., Park H.C., Yoo H.S., Kang D.W., Jin S.D., Min H.K., Paek W.K., Lim J. DNA barcoding of fish, insects, and shellfish in Korea. Genomics & informatics.2012; 10(3):206–211.
- Elizabeth L, Clare Æ, Kevin CR, Kerr Æ, Taika E, von Ko¨nigslo¨w Æ, John J, Wilson Æ, Paul DN, Hebert. Diagnosing Mitochondrial DNA Diversity: Applications of a Sentinel Gene Approach. J Mol Evol. 2008; 66:362-367.
- Brothers D.J. Phylogeny and evolution of wasps, ants and bees (Hymenoptera, Chrysidoidea, Vespoidea and Apoidea). Zoologica Scripta. 1999; 28: 233-250.
- Pilgrim E.M., Von Dohlen C.D., Pitts J.P. Molecular phylogenetics of Vespoidea indicate paraphyly of the superfamily and novel relationships of its component families and subfamilies. Zoologica Scripta. 2008; 37:539-560.
- Namin S.M., and Jury C. Genetic diversity of genus Vespa including an invaded species of V velutina (Hymenoptera: Vespoidea) in Korea inferred from DNA barcoding data. Journal of Asia-Pacific Entomology. 2020; 23: 540-545.
- Guz N., Kocak E. and Kilner N. Molecular phylogeny of Trissolcus species (Hymenoptera: Scelionidae). Biochemical systematics and ecology. 2013; 48: 85-91.
- Tang H., Jiang J., Wu T., Deng J. and Peng L. Six new species of Zaischnopsis Ashmead (Hymenoptera: Chalcidoidea: Eupelmidae) from China based on morphological and molecular data. European Journal of Taxonomy. 2022; 847: 1-27.
- Sohn J., Cornelis V.A., Yu Y. and Kim H. Two species new to South Korea, in the genus Apronopa Van Achterberg (Hymenoptera : Braconidae: Alysiine). Journal of Asia-Pacific Biodiversity. 2021; 14: 196-199.
- Venkatesan T., More R.P., Baskar R., Jalali S.K., Lalitha Y. and Ballal C.R. Differentiation of some indigenous and exotic trichogrammatids (Hymenoptera: Trichogrammatidae) from India based on Internal transcribed spacer-2 and cytochrome oxidase -1 markers and their phylogenetic relationship. Biological Control. 2016; 101:130-137.
- Zhou Q-S., Xi Y-Q., Yu F., Zhang X., Li X-J., Liu C-L., Niu Z-Q., Zhu C-D., Qiao G-X.and Zhang Y-Z. Application of DNA barcoding to the identification of Hymenoptera parasitoids from the soybean aphid (Aphis glycines) in China. Insect Science. 2014; 21: 363-373.
- Abd-El-Samie E.M., Elkafrawy I., Osama M. and Ageez A. Molecular phylogeny and identification of the Egyptian wasps (Hymenoptera: Vespidae) based on COI mitochondrial gene sequences. Egyptian Journal of Biological Pest Control. 2018; 28: 36.
- Chen H.Y., Li H.L., Pang H., Zhu C.D., and Zhang Y.Z. Investing the parasitoid community associated with the Invasive Mealybug Phenacoccus solenopsis in Southern China. Insects. 2021; 12:290.
- Parslow B.A., Schwarz M.P. and Stevens M.I. Molecular diversity and species delimitation in the family Gasteruptiidae. Genoma. 2021; 64: 253-264.
- Benavides A.L.P, Hernandez-Baz F., Gonzalez J.M., Romero-Napoles J., Hanson P.E. and Zaldivar-Riveron A. Integrative taxonomy to assess the species richness of Chalcidoid parasitoids (Hymenoptera) associated to Bruchinae (Coleoptera: Chrysomdidae) from Mexico. Revista Mexicana de Biodiversidad. 2020; 91:e913492.
- Siddiqui J.A., Chen Z., Li Q., Deng J., Lin X. and Hung X. DNA barcoding of aphid-associated ants (Hymenoptera, Formicidae) in a subtropical area of southern China. Zookeys. 2019; 879:117-136.
- Kwon Y.J., Lee M.L., Suh S.J. Phylogenetic relationship of Osmia species in Korea Based on mitochondrial COI gene sequences (Hymenoptera: Megachilidae). Journal of Asia-Pacific Entomology. 2003; 6(1): 7-14.
- Dowton M. and Austin A.D. Simultaneous analysis of 16S, 28S, COI and morphology in the Hymenoptera: Apocrita- evolutionary transitions among parasitic wasps. Biological Journal of the Linnean Society. 2008; 74(1):87-111.
- Aman-Zuki A., Mohammed M.A., Md-Zain B.M. and Yaakop S. Phylogenetic relationships of five oriental Apanteles species groups (Hymenoptera: Braconidae: Microgastrine) by concatenating four molecular markers. Journal of Asia-Pacific Entomology. 2019; 22:341-352.
- Niehuis O. and Waagele J.W. Phylogenetic analysis of the mitochondrial genes LSU rRNA and COI suggest early adaptive differentiation of anal teeth in Chrysidine cuckoo wasps (Hymenoptera: chrysididae). Molecular phylogenetics and Evolution. 2004; 30(3): 615-622.
- Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. DNA primers for amplification of mitochondrial Cytochrome Oxidase subunit I from diverse metazoan invertebrates. Molecular Biology. 1994; 3:294-299.

This work is licensed under a Creative Commons Attribution 4.0 International License.



