

How to Cite | Publication History | PlumX Article Matrix
P. Vanathi1* , Uma Sundaresan2 and Kavitha Manivannan3
, Uma Sundaresan2 and Kavitha Manivannan3
1Department of Microbiology, Bharathidasan Arts and Science College, Ellispettai, Erode, TamilNadu,India.
2Department of Food Process Engineering, School of Bioengineering, SRM University, Kattankulathur, Tamil Nadu, India.
3Department of Microbiology, Tharb Camel Hospital, Qatar.
Corresponding Author E-mail:vanathisekar3@gmail.com
DOI : http://dx.doi.org/10.13005/bbra/3325

ABSTRACT: Actinomycetes from mangrove and coastal environments were studied for their potential in biotechnology and environmental management. This research aimed to isolate and characterize these microbes, focusing on enzyme production, dye degradation, and antibiotic resistance. Samples were collected from various mangrove and coastal sites. Actinomycetes were isolated using selective media and identified through morphological and biochemical tests. Genetic characterization was performed using 16S rRNA sequencing. Enzyme production was evaluated through specific proteases, lipases, and cellulase assays. Dye degradation studies involved incubating actinomycetes with synthetic dyes and measuring degradation efficiency using spectrophotometric methods. Antibiotic resistance was assessed using disk diffusion and minimum inhibitory concentration (MIC) test. The study focused on isolating a variety of Actinomycetes from mangrove and coastal environments, assessing their potential for enzyme production and dye degradation.
KEYWORDS: Actinomycetes; Calcium carbonate; Dye degradation; 16SR RNA; SDS-PAGE
Download this article as:| Copy the following to cite this article: Vanathi P, Sundaresan U, Manivannan K. Characterization and Genetic analysis of Actinomycetes from Mangrove and Coastal Environments: Enzyme Production, Dye Degradation and Antibiotic Resistance. Biotech Res Asia 2024;21(4). |
| Copy the following to cite this URL: Vanathi P, Sundaresan U, Manivannan K. Characterization and Genetic analysis of Actinomycetes from Mangrove and Coastal Environments: Enzyme Production, Dye Degradation and Antibiotic Resistance. Biotech Res Asia 2024;21(4). Available from: https://bit.ly/48pZraI |
Introduction
Microorganisms can convert various raw materials into value-added products such as antibiotics, polysaccharides, proteins, oils, fatty acids, enzymes, and pigments. The synthesis of bioactive chemicals, industry, bioleaching techniques, pest and disease control, food production, and immunizations all depend on a diverse range of microorganisms1,2. Actinomycetes, filamentous, free-living, soil-aerobic bacteria, play a specific role in the production of antibiotics and other secondary metabolic products3. They fall into three categories: auto regulators, agro-biological active agents, and pharmacological active agents4.
Actinomycetes are a diverse group of gram positive bacteria known for their complex life cycles and ability to produce a variety of bioactive compounds. These microorganisms are abundant in soil environments and are particularly notable in extreme and niche habitats, such as mangrove and coastal environments. Because of their unique sediment composition and saline environment, Actinomycetes are abundant in mangrove ecosystems and coastal habitats and have potential biotechnological applications5, 6. About 2/3 of naturally occurring antibiotics have been isolated from these organisms. Streptomyces, the largest antibiotic-producing genera in the microbial world, generate an extensive range of secondary metabolites with important applications in pharmaceutical industries7.
In recent years, Interest has been rising in exploring Actinomycetes from these habitats for their unique metabolic capabilities. Actinomycetes have been shown to produce a range of enzymes, including laccases, which are of significant industrial interest due to their applications in bioremediation and the degradation of recalcitrant dyes8. Laccases are oxidoreductases that can degrade an assortment of environmental pollutants, making them valuable in wastewater treatment processes9.
Dye degradation involves the cleavage of chromophores, with laccases being the most promising enzymes10. They are effective in oxidative elimination of aromatic alcohols and decolorization of effluents. Gene manipulation, particularly protoplast fusion, can investigate the importance of genetic traits in bio degradative compounds11. Actinomycetes, less pathogenic than bacteria and fungi, are a good source of useful enzymes for dye oxidation. Due to stringent environmental legislation, textile industries are looking for effective wastewater re-mediation technologies for color removal12-14. Bioremediation of textile effluent is considered an inexpensive, environmentally friendly, and publicly acceptable treatment technology 15. Protoplast fusion technology can achieve a high decolorizing capacity of actinomycetes, reducing dye.
Actinomycetes’ capacity to break down dyes and other contaminants is associated with their enzymatic systems, which are distinguished by the generation of color and the activity of enzyme hydrolysis 16. The presence of antibiotic-resistant microorganisms in environmental samples presents both opportunities and concerns. Investigating these strains aids in the development of novel antibacterial tactics as well as our understanding of the mechanisms driving antibiotic resistance17.
The study focuses on the deterioration of dyes by Laccase enzyme-producing Actinomycetes, as industries often discharge toxic dye effluents into water bodies, posing significant environmental and human health risks.
Materials and Methods
Collection of Samples
In all, 50 mangrove soils, 70 beach littoral sediments, 70 beach water samples, and about 75 coir waste samples were collected in sterile containers. At a depth of 10–15 cm, mangrove soil and beach littoral sediment were collected. All the above samples were collected from various regions of Kerala and Tamilnadu and were brought to the laboratory for further processing.
Isolation of Actinomycetes
Actinomycetes were isolated from the samples by both pretreatment and without pretreatment methods. The pretreatment methods used were (i) Calcium carbonate treatment 18and (ii) Oven treatment of the sample at 55°C for 6 min in a hot air oven 19. Then the samples were serially diluted and plated on starch casein nitrate agar20, 21 with cycloheximide and nystatin, each at 50 g/mL, and rifampicin at 0.5 mL22,23. The plates were incubated at 27°C for 7–10 days. After incubation, the colonies formed were taken for further study. Each isolate was selected based on the nature of the colony, the nature of the odor, and also the colour of the pigment diffused into the medium. They were named accordingly as S1, S2, S3, S4, S5, etc.
Gram Staining
Gram staining was performed for all the strains using a sterile technique, and the results were tabulated24.
Melanoid Pigment Production
Melanoid pigment production was observed on yeast extract malt extract agar. The cultures forming colours, from greenish brown to brown or black diffusible pigment or a distinct brown pigment modified by other colours, should have to be recorded. The absence of any of these colours was considered negative for pigment production.
Carbohydrate Fermentation Tests
A carbohydrate fermentation test was performed to detect the fermentation activity of the actinomycetes25. Using various carbon sources, each test culture was inoculated into the sterile broth used for the fermentation of carbohydrates (glucose, sucrose, fructose, raffinose, inositol, xylose, and mannitol) and incubated at 27°C for 7-10 days.
Hydrolytic Activity of the Isolates
The enzymatic potential such as amylase, gelatinase and caseinaseactivity of the isolates was tested using starch agar, gelatine agar and skim milk agar26.
Determination of Laccase Enzyme Activity
For the determination of laccase enzyme activity, both starch casein nitrate agar medium together with guaiacol (1 mM) and peptone with guaiacol (1 mM) were employed20. The cultures were cultured at 27°C for 7–10 days after being inoculatedinto the plates using the direct streaking method. After incubation, the plates were inspected for the presence or absence of zones that indicated either hydrolysis or inhibition. In particular, zones of hydrolysis were analyzed to detect enzyme activity, whereas zones of inhibition were assessed to evaluate antimicrobial effects.
Partial Purification of the Laccase Enzyme
The partially purified form of the enzyme obtained was undergone for molecular weight determination27.
Confirmation of Laccase Enzyme by SDS-PAGE Method
SDS-PAGE was used to estimate the molecular weight of the laccase enzyme 28. SDS-PAGE was done to estimate the molecular weight of the laccase enzyme using Laemmli’s method. The running buffer contained 0.125 M Tris, 0.96 M glycine, and 0.5% SDS. Separating gel consisted of 1.5 M TrisHCl, 0.4% SDS, and 30% acrylamide, among others, while stacking gel had 0.5 M TrisHCl, 0.4% SDS, 30% acrylamide, etc. Samples with tracking dye and mercaptoethanol were loaded into wells, and electrophoresis was carried out at 50V. The staining solution included methanol, acetic acid, and Coomassie Brilliant Blue R. The gel was stained overnight, washed, and then destained in another solution.
Isolation of DNA, Amplification and Sequencing of 16SrRNA Gene
Broth culture was used to extract the DNA, with 10% SDS and 1 X TE buffer. DNA was extracted by adding phenol, chloroform, isoamyl alcohol, and then isopropanol. The DNA fragments were then removed by centrifuging the mixture, and Electrophoresis was performed using 1% agarose gel. Genomic DNA was subjected to PCR assays for amplification of the 16SrRNA gene. PCR assays utilized 29 to amplify the 16S rRNA gene, determining partial and full-length gene sequences accessible. Primers from Sigma in India, Primer 1 (5′-GAGAGTTTGATCCTGGCT-3′) and Primer 2 (5′-TGACGGGCGGTGTGTA-3′), produced a 1400 bp amplicon. Each 20 μl PCR reaction mixture contained 1 μl genomic DNA template, 2 μl 10X PCR buffer, 0.5 μl of each primer, 1 μl of each dNTP, 0.5 μlTaq DNA polymerase, and 15 μl water. Samples underwent 35 cycles of denaturation at 94°C for 30 seconds, annealing at 63°C for 30 seconds, and extension at 72°C for 30 seconds. Gel analysis with ethidium bromide revealed the amplification band for evolutionary relationship study.
Dye Degradation of Actinomycetes
Positive laccase enzyme cultures were tested for dye degrading efficiency in starch casein nitrate broth with various dyes with different concentrations. After 7-10 days of incubation at 27°C, culture samples were analyzed for decolorization progress by monitoring absorbance at specific wavelengths and pH changes. Decolorization percentage was calculated based on the results of the study.Decolorization was monitored by measuring the absorbance of culture supernatants at respective nm. pH change during decolorization was observed to see whether decolorization was due to degradation or change in pH. The percentage decolorization was calculated as follows,

The results were tabulated.
Antibiotic Resistant Mutant Isolation
In an experiment by different concentrations of antibiotics 30 were added to starch casein nitrate agar medium to identify antibiotic-resistant bacteria. Plates were then incubated at 27°C for 7-10 days, resulting in growth of actinomycetes being observed.
Protoplast Fusion
The conducted protoplast fusion using the S9 (Str+R-) and S19 (Str-R+) strains31. Protoplasts were produced by treating mycelia with lysozyme, rinsing, and incubating them on agar plates. Protoplast lysate preparation involved centrifuging, osmotically breaking, and mixing with sucrose before incubation. Adding donor DNA, PEG, and antibiotics before plating are essential for growth observation in the transformation process. Protoplast fusion allows genetic material merging from different strains, potentially leading to beneficial traits in hybrids. Agar plate growth monitoring demonstrated successful gene transfer and integration evaluation.
Dye Degradation of Fused Strains
The efficiency of dye degradation by fused strains was tested using different dyes in starch casein nitrate broth. After incubation at 27°C for 7-10 days, samples were taken at intervals and centrifuged. Decolorization was measured by absorbance at specific wavelengths, with pH changes noted. The percentage of decolorization was calculated to determine the effectiveness of the strains in degrading the dyes.
Decolorization was monitored by measuring the absorbance of culture supernatants at respective nm. pH change during decolorization was observed to see whether decolorization was due to degradation or change in pH. The percentage decolourization was calculated as follows,

The results were tabulated.
Results
Isolation of Actinomycetes
Samples were obtained from different locations in Kerala and Tamilnadu, including 50 wetland soils from mangrove forests, 70 sediments from beach shores, 70 beach water samples, and 75 coconut waste samples. The samples were subjected to both pre-treatment and left in their untreated state.The use of calcium carbonate pre-treatment was shown to be more effective for isolating actinomycetes compared to other methods utilized (Table 1).
Table 1: Number of Colonies of Actinomycetes in Different Sampling Station
| Samples | Station | Number Of Colonies Cfu/Ml 10-2 | ||
| Calcium Carbonate Treatment | Oven Treatment | Without Treatment | ||
| Mangrove Soil | Ochira | 20 | 8 | 15 |
| Kumarakom | 25 | – | 12 | |
| Beach Sediment | Ambalapuzha | 20 | 2 | 15 |
| Kovalam | – | – | 5 | |
| Cherthala | 12 | 2 | 6 | |
| Ernakulam | 11 | – | 8 | |
| Kollam | – | – | – | |
|
Beach Water |
Ambalapuzha | 9 | 4 | 6 |
| Kovalam | 5 | – | 2 | |
| Cherthala | 2 | – | 1 | |
| Ernakulam | 4 | 2 | 2 | |
| Kollam | – | – | – | |
| Coir Waste | Kollam (A) | 62 | – | 30 |
| Kollam (B) | 40 | – | – | |
| Vijayamangalam (A) | 38 | – | 14 | |
| Vijayamangalam (B) | 45 | 1 | 8 | |
Table 2: Colony Morphological Characteristics of the Isolates
| S.No. | Strain | Sample | Colony Morphology
|
| 1. | S2 | Mangrove Soil | Small circular, white powdery colonies with brown pigmentation. |
| 2. | S6 | Mangrove Soil | Small, white powdery colonies with grayish circle. |
| 3. | S9 | Coir Waste | White powdery, raised mucoid colony with white colour on reverse side. |
| 4. | S12 | Mangrove Soil | Small, circular white colony with pointed centre. |
| 5. | S18 | Beach Sediment | Circular grayish powder colonies. |
| 6. | S19 | Coir Waste | White, mucoid, powdery colony with raised centre. |
| 7. | S25 | Coir Waste | Brown coloured colony with sandal colour on reverse side. |
| 8. | S26 | Coir Waste | Grey colored colony raised at centre with reddish brown pigmentation on reverse side. |
| 9. | S28 | Coir Waste | Large circular yellow coloured powdery colony. |
| 10. | S109 | Beach Sediment | Large circular, grayish white powdery colonies with green pigmentation. |
| 11. | S110 | Mangrove Soil | Large, white powdery colonies with yellow pigmentation. |
| 12. | S111 | Mangrove Soil | Large circular, grayish powdery colonies with black pigmentatiom. |
The colonies were selected randomly according to their morphology, odour, powdery nature and pigmentation and were named as S1, S2, S3,………S111 (Table.1, Plate: 1).
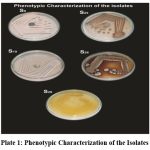 |
Plate 1: Phenotypic Characterization of the Isolates |
All isolates listed (S2, S6, S9, S12, S18, S19, S25, S26, S28, S109, S110, S111) show a positive result for Gram staining. This indicates that every strain is Gram-positive, which implies that they still display the crystal violet stain and have a thicker coating of peptidoglycan in their cell walls. Only one strain (S26) of the 12 isolates exhibits positive melanoid pigment synthesis. . This suggests that strain S26 produces melanoid pigments. While most of the strains do not. Melanoid pigments are typically brown to black and can be produced by certain bacteria through the breakdown of amino acids or carbohydrates, and their production is often associated with specific metabolic pathways.
Table 3: Utilization of the Carbon Sources by the Isolates
| S.No. | Carbohydrate (Sugars)
|
Actinomycetes Strains
|
|||||||||||
| S2 | S6 | S9 | S12 | S18 | S19 | S25 | S26 | S28 | S109 | S110 | S111 | ||
| 1. | Glucose | – | – | + | + | – | – | – | – | – | – | – | – |
| 2. | Fructose | + | + | + | + | + | + | + | + | + | + | + | + |
| 3. | Sucrose | + | – | – | + | – | – | + | – | – | – | – | – |
| 4. | Xylose | – | – | + | + | + | + | – | – | + | + | – | + |
| 5. | Mannitol | – | + | – | – | – | – | – | + | – | – | – | – |
| 6. | Raffinose | – | – | – | – | – | – | – | – | – | – | – | – |
| 7. | Inositol | – | – | – | – | – | – | – | – | – | – | – | – |
(+) – positive; -(-), Negative
In carbohydrate fermentation, the strains showed variation in their ability to assimilate seven carbon sources; none of the strains grew on substrates like inositol and raffinose(Table. 3). All strains (S2, S6, S9, S12, S18, S19, S25, S26, S28, S109, S110, S111) can utilize fructose, indicating it is a common and essential carbohydrate source for these strains. The strains S9, S12, S18, S19, S28, S109, and S111 fermented xylose, and others could not. Only strains S9 and S12 fermented glucose, and strains S6 and S26 fermented mannitol. All the isolates fermented fructose, and the strains S2, S9, S12, and S25 fermented sucrose.
Table 4: Isolates Showing Various Hydrolytic Enzyme Activities
| S.No. | Strain | Starch Hydrolysis | Casein Hydrolysis | Gelatin Hydrolysis |
| 1. | S2 | + | + | – |
| 2. | S6 | + | + | + |
| 3. | S9 | + | – | + |
| 4. | S12 | + | + | + |
| 5. | S18 | – | – | – |
| 6. | S19 | + | + | + |
| 7. | S25 | – | + | – |
| 8. | S26 | + | – | + |
| 9. | S28 | + | – | + |
| 10. | S109 | + | + | + |
| 11. | S110 | + | + | + |
| 12. | S111 | + | – | + |
(+)- Positive; (-)- Negative
The enzymes hydrolysis (amylase, caseinase, and gelatinase) were tested for all the strains(Table.4). A majority of strains (S2, S6, S12, S19, S25, S109, S110) can hydrolyze casein, showing they can degrade casein into peptides and amino acids. Strains S9, S18, S26, S28, and S111 do not show casein hydrolysis. Several strains (S6, S9, S12, S19, S26, S28, S109, S110, and S111) can hydrolyse gelatin, indicating they have the enzyme gelatinase, which breaks down gelatin into simpler products. Strains S2, S18, and S25 cannot hydrolyzegelatin. The strains S2, S6, S9, S12, S19, S26, S28, S109, S110, and S111 were capable of hydrolyzing starch, which showed a positive result for amylase, and the strains S18 and S25 showed negative results (Plate: 2).
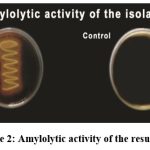 |
Plate 2: Amylolytic activity of the results |
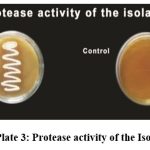 |
Plate 3: Protease activity of the Isolates |
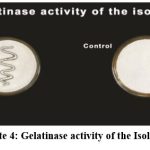 |
Plate 4: Gelatinase activity of the Isolates |
The strains S2, S6, S12, S19, S25, S109 and S110 showed positive result for casein and the strains S9, S18, S26, S28 and S111 showed negative result (Plate: 3). The strains S6, S9, S12, S19, S26, S28, S109, S110 and S111 were found to be positive for gelatin and the strains S2, S18 and S25 showed negative result ( Plate:4).
The laccase enzyme assay was carried out by using both starch casein nitrate agar with guaiacol and peptone with guaiacol. Strains S2, S6, S12, S18, S109, S110, and S111 do not show positive results for any of the tests, suggesting limited or different metabolic activities compared to the others. The strains S9, S19, S25, S26 and S28 showed reddish brown zones on starch casein nitrate agar with guaiacol. In the peptone with guaiacol medium greenish fluorescent culture growth was observed ( Plate: 5).
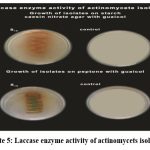 |
Plate 5: Laccase enzyme activity of actinomycets isolates |
The laccase enzyme was partially purified and the SDS-PAGE was performed for confirmation of laccase which showed the molecular weight of the laccase enzyme is nearing ~35 KDa to ~75 KDa (Plate: 6).
 |
Plate 6: Protein Profile of the Isolates (SDS-PAGE) |
The isolate was screened for DNA isolation and a band was observed. The molecular weight of the isolated strain was 1400 bp (Plate: 7).
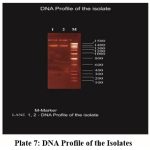 |
Plate 7: DNA Profile of the Isolates |
Isolation of DNA, Amplification and Sequencing
The phylogenetic analysis of 16Sr RNA confirmed that the isolate showed similarities of the genus Actinomyces(Fig. 1-3).
 |
Figure 1: Phylogram |
 |
Figure 2: Cladogram |
 |
Figure 3: Phylo Draw Results |
Dye degradation
The isolates that were laccase-positive were selected for the dye decolorization. The azo dyes chosen were reactive dye N-blue M2R, direct dye Swiss pink, and orange ME2RL. Throughout the procedure, the absorbance and pH were checked every 24 hours. The absorbance was measured to determine the percentage of decolorization. Strain S9 exhibited the highest percentage of decolorization (66%) for both the reactive dye N-blue M2R and the direct dye Swiss Pink (86%), while strain S28 displayed the highest percentage (52%) for orange ME2RL after 48 hours in a 50µl dye concentration of all the isolates, these results were obtained(Fig: 4-6).
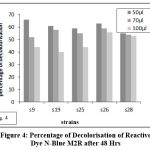 |
Figure 4: Percentage of Decolorisation of Reactive Dye N-Blue M2R after 48 Hrs |
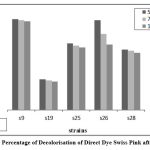 |
Figure 5: Percentage of Decolorisation of Direct Dye Swiss Pink after 48 Hrs |
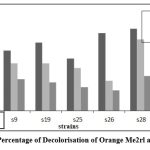 |
Figure 6: Percentage of Decolorisation of Orange Me2rl after 48 Hrs |
After 120 hours, strain S9 showed higher percentage of decolourization for reactive dye N- blue M2R (86%) and direct dye swiss pink (89%) and S25 showed higher percentage of decolourization for orange ME2RL (70%) in 50µl concentration of the dye (Fig: 7-9). The highest percentage of decolourization was observed in 50µl concentrations of the dye compared to 70µl and 100µl of dye concentration.
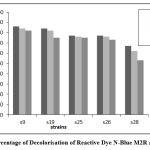 |
Figure 7: Percentage of Decolorisation of Reactive Dye N-Blue M2R after 120 Hrs |
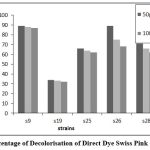 |
Figure 8: Percentage of Decolorisation of Direct Dye Swiss Pink after 120 Hrs |
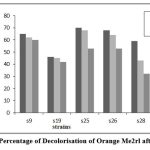 |
Figure 9: Percentage of Decolorisation of Orange Me2rl after 120 Hrs |
All the strains were checked for one step and multi-step resistant mutant for the following antibiotics streptomycin, rifampicin, gentamycin, amoxycillin and chloramphenicol. All the strains were resistant to gentamycin, chloramphenicol, and amoxycillin at all concentrations. The strain S19 showed sensitivity to 100 ul of streptomycin concentration, and other strains S9, S25, S26, and S28 showed resistance to all the concentrations of streptomycin. The strain S9 showed sensitivity to 50 ul of rifampicin, and strain S19 showed resistance to 50 ul of rifampicin concentration. In the concentration of 800 ul, S25 showed sensitivity, while S26 showed sensitivity from the 500 ul concentration, and S28 showed sensitivity from the 600 ul concentration.The strains S9 and S19 were taken for the protoplast fusion according to the antibiotic mutant pattern (S9R-Str+ x S19R+Str-). S9 was chosen for protoplast formation; after protoplast formation, strain S9 was checked for regeneration on starch casein nitrate agar and regeneration medium. The growth was observed.
Following the preparation of the protoplast lysate, growth observation was also performed on strain S19. Since no growth was seen, more actions were taken. To construct the protoplast and protoplast lysate (S9 x S19) transformation, polyethylene glycol (PEG) 4000 was utilized. Following protoplast fusion, the merged strains were grown on starch casein nitrate agar and R medium, which has 50 ul of rifampicin and 100 ul of streptomycin. It was noted that growth occurred in both media. After being plated on the medium, the control (which has no PEG 4000 and the other has no lysate) showed no signs of growth on either plate.
The fused strains (S9 x S19) were checked for dye decolorization, and the percentage of dye decolorization was calculated. In 24 hours, the fused strains in SCNA medium showed a higher percentage of decolorization for reactive dye N-blue M2R (21%), direct dye Swiss pink (14%), and orange ME2RL (21%) in 50 l dye concentration. After 48 hours, the fused strain in starch casein nitrate agar showed a higher percentage of decolorization for reactive dye N-blue M2R (73%), direct dye Swiss pink (68%), and for the dye orange ME2RL (53%), in 50 ul dye concentration (Fig.10-13). The fused strains in the SCNA medium showed a higher percentage of decolorization than the fused strains in the R medium. Additionally, at concentrations of 70 and 100 ul, there was decolorization, but the proportion of decolorization was lower than at the 50 ul concentration.
 |
Figure 10: Fused strain- Percentage of Decolorisation of Reactive Dye N-Blue M2r after 24 Hrs |
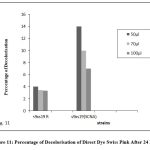 |
Figure 11: Percentage of Decolorisation of Direct Dye Swiss Pink After 24 Hrs |
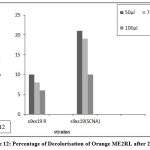 |
Figure 12: Percentage of Decolorisation of Orange ME2RL after 24 Hrs |
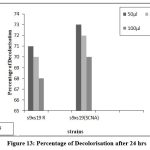 |
Figure 13: Percentage of Decolorisation after24hrs |
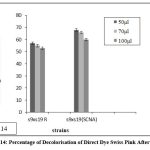 |
Figure 14: Percentage of Decolorisation of Direct Dye Swiss Pink After 48 Hrs |
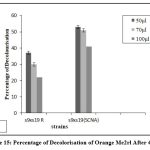 |
Figure 15: Percentage of Decolorisation of Orange Me2rl After 48 Hrs |
Discussion
In recent years, there has been a growing interest in rare actinomycetes from uncharted regions as potential sources of new biotic compounds to facilitate genetic analysis, molecular breeding, and the extraction of these organisms for the breakdown of enduring compounds, and an effective transformation system for rare actinomycetes is necessary. Therefore, the study focused on the ability of protoplast-fused actinomycetes that produce laccase enzymes to degrade dyes. Both the pretreatment and untreatment methods were utilized to isolate actinomycetes from coir waste, mangrove soil, littoral sediment, and beach water samples. Actinomycetes were successfully recovered at a rate of 81% from the overall microbial population using the calcium carbonate method, as demonstrated in the work of19, 21, 22, which supports its application.
Their statement timely validated our investigation, as more colonies could only be isolated when the calcium carbonate pretreatment method was applied. Originally, pretreatment methods were not appropriate for the isolation. They used the oven method at 55°C for 15 minutes and 100°C for 60 minutes. Because fewer colonies were formed using the oven treatment method than with calcium carbonate, it was shown to be ineffective for isolating actinomycetes in the current investigation 32,33.
The isolation process in the current study utilized starch casein nitrate agar with cycloheximide, nystatin, and rifampicin, following the same procedure as previous studies34,35. Gram staining results revealed the presence of gram-positive actinomycetes. In the current research, all the isolates were also found to be gram-positive. Only a small number of actinomycetes genera produce a melanin pigment that can spread, whereas, in the current research, only one strain has been observed to generate such a pigment 36-38.
During the carbon utilization experiments, abundant growth of the actinomycete colonies indicated that the isolates efficiently used the specific carbon source. The test results were positive for fructose, meso-inositol, mannitol, and glucose but negative for xylose, raffinose, and sucrose. In this study, all strains tested were found to be positive for fructose and negative for raffinose, with only a small number of strains fermenting glucose and mannitol, consistent with previous research. The isolates tested negative for inositol, while some strains fermented sucrose and xylose, showing different results. A small number of isolates were able to hydrolyze starch and gelatin, while a few strains were able to hydrolyze starch and casein. 80% of these isolates exhibited activity in amylase, caseinase, and gelatinase, which prompted our current research. In the current research, the majority of the strains tested positive for starch, gelatin, and casein.
In this study, 39 a combination of 0.04% remazol brilliant blue R (RBBR), 0.04% polymeric dye R-478, 0.01% guaiacol, and 0.5% tannic acid employed to identify laccase enzyme 39. Out of these indicators, guaiacol demonstrated more favorable outcomes by yielding a brown hue. 40 Polymeric dye R-478, remazol brilliant blue R, cinnamic acid, vanillic acid, ferulic acid, and guaiacol was used in an agar medium with peptone and glucose for laccase assay40. A clear zone around colonies was observed when guaiacol was present at a concentration of 1 mM. In this study, two agar mediums were utilized, with strains showing a reddish brown zone in starch casein nitrate agar with guaiacol and appearing fluorescent green in peptone guaiacol media without zone formation. Starch casein nitrate agar medium with guaiacol was found to be the most effective in screening laccase assay among the two agar mediums 41. The laccase enzyme assay performed on strains from coir waste exhibited superior results when compared to those from mangrove soil and beach samples42.
The isolated protein had a molecular weight of about 35 KDa, which fell within the range of 32 KDa to 75 KDa for other proteins43,26. The dimeric form of laccase from Streptomyces ipomoea has a molecular mass of 79 KDa, while the monomeric form has a mass of 44.7 KDa23.The DNA molecular weight of actinomycetes ranges from 600 bp to 1400 bp44,45 . In the current research, the actinomycetes DNA had a molecular weight of about 1400 bp, which was confirmed. The analysis of the isolated strains’ evolutionary relationships indicated a resemblance to the genus Actinomyces, by mirroring findings 46.
In a study, it was found that S. krainskii SUK-5 achieved 95% and 87% decolorization of Navy blue RX at 30-50 mg/l47. However, doses of 70–100 mg/l were found to be harmful to the organism. The complete decolorization of the dye took around 24 and 48 hours for 30 and 50 mg/l doses, respectively. Azo dyes experienced a lower level of decolorization at higher concentration levels compared to lower levels due to the lethal impact of the higher concentration. The same pattern was noticed in the current investigation. In previous research, the most significant color removal was observed within 2 to 3 days, whereas in our current study, it took 3 to 5 days to achieve the greatest color removal of approximately 89%48. The research provided clear guidance for selecting the streptomycin and rifampicin-resistant and sensitive strains for the protoplast fusion study (S9 R-Str+ x S19 R+Str-)49
It was observed the development 50of the Streptomyces parvulus protoplast on the recovery medium within 5 to 10 days 50. This analysis revealed colonies on the regeneration medium and starch casein nitrate agar medium after 6 days. The researchers noticed a change when polyethylene glycol was present, and they carried out their transformation experiments according to their established methods. Protoplasts and genetically altered cells were cultivated on R medium and starch casein nitrate agar in this investigation. Growth was visible on both media after 4 days. The successful inoculation on R medium, followed by cloning the laccase gene51 from S. coelicolar into S. lividans, resulting in efficient decolorization of indigo dye by the purified laccase. Within a 20-minute incubation period at 45°C, more than 85% decolorization was achieved for the dye. Using recombinant laccase, along with a redox mediator and acetosyringone, led to a 90% reduction indecolorization of azo dye orange-II after 4 hours of incubation in various studies52-54. In the current research, decolorization began after 24 hours, with a 70% decolorization rate achieved within 48 hours by fused strains (cultivated on starch casein nitrate agar). The fused strain grown on starch casein nitrate agar medium exhibited a greater proportion of discoloration in comparison to the fused strain grown on R medium. Out of the samples analyzed in this study, the coir waste, the ability of isolates to produce laccase enzyme and decolorize textile dyes was good. The above details have provided improved satisfaction for continued education.
Conclusion
Actinomycetes were found in mangrove and coastal environments, displaying diverse characteristics and strong enzyme-producing abilities, particularly for proteases, lipases, and cellulases. This suggests their potential in various industrial applications, such as waste management and bio-transformation. Some strains showed efficient dye degradation capabilities, highlighting their potential in bioremediation for treating dye-contaminated wastewater. Different strains exhibited varying efficiency in dye degradation, emphasizing the need for further research. Certain strains also displayed antibiotic resistance, emphasizing the importance of managing resistance in environmental microorganisms. Future research should focus on optimizing enzyme synthesis and dye breakdown processes, as well as understanding genetic mechanisms behind antibiotic resistance, to develop effective solutions. Exploring additional applications, such as pharmaceuticals and advanced bioremediation, could unlock more benefits from these microbes.
Acknowledgment
The Department of Microbiology at Bharathidasan College of Arts & Science is commended for its comprehensive support and collaborative teamwork in this publication.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Authors’ Contributions
P.V and U.S (P.Vanathi & Uma Sundaresan): Resources, Conceptualization, Methodology, Writing – Original Draft.
U.S (Uma Sundaresan): Analysis, Writing – Review & Editing.
P.V and K.M. (P.Vanathi & Kavitha Manivannan): Visualization, Supervision, Project Administration
References
- Khan MI, Singh A, Kumar P.Microbial biotechnology: Recent advances and applications. ApplMicrobiolBiotechnol.2022; 106(3): 1039-1051.
- Singh R, Verma S, Gupta R.Innovative approaches in bioactive compound synthesis. J Biotechnol. 2023; 348: 1-15.
- Zhang Y, Chen Y, Wang H. (2021). Role of Actinomycetes in antibiotic production. Front Microbiol.2021; 12: 645-658.
- Patel A, Kumari P. Classification and applications of Actinomycetes. Current Trends in Microbiology, 2022; 12(4): 317-329.
- Goodfellow M, Fida T, Tanaka Y. Actinomycetes in Marine Environments. In The Actinobacteria 2012; 2: 25-50.
- Madhusudhan P, Reddy KK, Kumar NS.Ecological significance and biotechnological potential of Actinomycetes in mangrove ecosystems.Micro Ecol. 2023; 86(1): 123-135.
- Thompson CC, Santos MV, Lima AL. Streptomyces: A treasure trove of bioactive compounds. J Antibiotic. 2022; 75(4): 233-250.
- Kumar CG, Hsu SH. Laccase Enzymes: Properties and Applications. J Biotech. 2021; 123(4):589-600.
- Sánchez C, Romero A. Biotechnological Applications of Laccase. Biotechnol Adv. 2017; 35(5): 705-715.
- Ngo ACR, Tischler D. Microbial Degradation of Azo Dyes: Approaches and Prospects for a Hazard-Free Conversion by Microorganisms. Int J Environ Res Public Health. 2022 Apr 14;19(8):4740.
CrossRef - Reed KM, Bargmann BOR. Protoplast Regeneration and Its Use in New Plant Breeding Technologies. Front Genome Ed. 2021; 3:734951.
CrossRef - Saini A, Aggarwal NK, Sharma A, Yadav A. Actinomycetes: A Source of Lignocellulolytic Enzymes. Enzyme Res. 2015; 1:279381.
CrossRef - Tripathi M, Singh S, Pathak S, Kasaudhan J, Mishra A, Bala S, Garg D, Singh R, Singh P, Singh PK, Shukla AK, Pathak N. Recent Strategies for the Remediation of Textile Dyes from Wastewater: A Systematic Review. Toxics. 2023; 11(11):940
CrossRef - Sharma J, Sharma S, Soni V. Classification and impact of synthetic textile dyes on Aquatic Flora: A review. Reg Stud Mar Sci. 2021 Jun 1; 45:101802.
CrossRef - Azanaw A, Birlie B, Teshome B, Jemberie M. Textile effluent treatment methods and eco-friendly resolution of textile wastewater. Case Studies Chem Environ Eng. 2022 Dec 1;6:100230.
CrossRef - El Awady ME, El-Shall FN, Mohamed GE, Abd‑Elaziz AM , Abdel‑Monem MO , Hassan Exploring the decolorization efficiency and biodegradation mechanisms of different functional textile azo dyes by Streptomyces albidoflavus 3MGH. BMC Microbiol .2024;24: 210 .
CrossRef - Gordon RE, Williams ST. Actinomycetes for Soil Bioremediation. Environ Sci Technol. 2019; 53(10):5891-5900.
- MartínezJL,Fajardo A. Antibiotic Resistance Mechanisms in Actinomycetes. ClinMicrobiol Rev. 2020; 33(1): e00032-19.
- Tsao PH, Leben C, Keitt GW. An enrichment method for isolating actinomycetes that produce diffusible antifungal antibiotics. Phytopathology. 1960;50: 88-89.
- Niladevi KN, Prema P. Mangrove actinomycetes as the source of ligninolytic enzymes. Actinomycetologica. 2005; 19(2): 40-47..
CrossRef - Rifaat R, Saadoun I, Mahasneh A. Effect of cultural conditions on xylanase production by Streptomyces species (Strain lb 24D) and its potential to utilize tomato pomace. Afr 2005; 4(3): 251-255.
- El-Nakeeb, MA, LechevalierHA.Selective isolation of aerobic actinomycetes. ApplMicrobiol. 1962; 11: 75-77.
CrossRef - Joe D’ Souza, Vaidya R. Screening estuarine actinomycetes for antibiotic production. Asian J MicrobiolBiotechnol Environ Sci.1999; 1(1-2): 47-52.
- Johnson LA, Smith RT, Lee CH. Techniques for the characterization of bacterial strains: A review. J Microbiol Methods, 2023; 204:106123
- Vasavada SH, Thumar JT, Singh SP. Secretion of a potent antibiotic by salt- tolerant and alkaliphilicactinomycete Streptomyces sannanensis strain RJT-1. Curr Sci.2006; 91(10): 1393-1397.
- Arias ME, Arenas, M, Rodriguez. J, Soliveri J, Ball AS, Hernandez M. Kraft pulp biobleaching and mediated oxidation of a nonphenolic substrate by laccase from Streptomyces cyaneus CECT 3335. Appl Environ Microbiol. 2003; 69: 1953-1958.
CrossRef - Laemmli, UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4.1970; 227: 680-685.
CrossRef - Mane UV, Gurav PN, Deshmukh AM, Govindwar SP. Degradation of textile dye reactive navy- blue Rx (Reactive blue-59) by an isolated actinomycetes Streptomyceskrainskii SUK-5. Malays J Microbiol.2008; 4(2): 1-5.
- Annette B, Smith J, Jones M. PCR amplification and sequencing of the 16S rRNA gene for bacterial identification.J Microbial Methods.2003; 54(2): 123-132.
- Lee DH, Miles RJ, Inal JRM. Antibiotic sensitivity and mutation rates to antibiotic resistance in Mycoplasma mycoides species mycoides. Epidem Inf. 1987; 98: 361-368.
CrossRef - Kozo O. Protoplast fusion permits High- frequency transfer of a Streptomyces determinant which mediates actinomycin synthesis. J Bacteriol. 1982; 150(2): 592-597.
CrossRef - Meenakshi S, Hiremath J, Meenakshi MH, Shivaveerakumar S. Actinomycetes: Isolation, Cultivation and its Active Biomolecules. J Pure ApplMicrobiol. 2024;18(1):118-143.
CrossRef - Tiwari D, Shouche S, Bhati P, Das P. A Consolidated Method for Selective Isolation of Actinomycetes Based on Choice of Substrate. Int Ann Science. 2021 Dec 11;11(1):10-21.
CrossRef - Rosmine E, Varghese SA. Isolation of actinomycetes from mangrove and estuarine sediments of Cochin and screening for antimicrobial activity. J Coast Life Med. 2016;4(3):207-10.
CrossRef - Subramani R, Sipkema D. Marine Rare Actinomycetes: A Promising Source of Structurally Diverse and Unique Novel Natural Products. Mar Drugs. 2019;17(5):249.
CrossRef - Abraham J, Chauhan R. Profiling of red pigment produced by Streptomyces JAR6 and its bioactivity. 3 Biotech. 2018; 8(1):22.
CrossRef - Sundar R, Sivaperumal P. Melanin pigments from sediment-associated Nocardiopsis sp. marine actinobacterium and antibacterial potential. J Adv Pharma Technol Res. 2022 Nov 1;13(Suppl 1):S88-92.
CrossRef - El-Zawawy NA, Kenawy ER, Ahmed S, El-Sapagh S. Bioproduction and optimization of newly characterized melanin pigment from Streptomyces djakartensis NSS-3 with its anticancer, antimicrobial, and radioprotective properties. Microb Cell Fact. 2024 Jan 17;23(1):23.
CrossRef - Kiiskinen, LL, Ratto M, Kruus, K. Screening for novel laccase producing microbes. J Appl Microbiol.2004; 97: 640-646.
CrossRef - Wikee S, Hatton J, Turbé-Doan A, Mathieu Y, Daou M, Lomascolo A, Kumar A, Lumyong S, Sciara G, Faulds CB, Eric R. Characterization and Dye Decolorization Potential of Two Laccases from the Marine-Derived Fungus Pestalotiopsissp. Int J Mol Sci. 2019; 20(8):1864.
CrossRef - Abu-Hussien SH, Hemdan BA, Alzahrani OM, Alswat AS, Alatawi FA, Alenezi MA, Darwish DBE, Bafhaid HS, Mahmoud SF, Ibrahim MFM, El-Sayed MS. Microbial Degradation, Spectral analysis and Toxicological Assessment of Malachite Green Dye by Streptomyces exfoliatus. Molecules. 2022; 27(19):6456.
CrossRef - Thrimothi D, Sujatha E, Swetha KG, Krishna G. Isolation, Screening, Identification, and Assessment of Laccase-Producing Fungi Isolated From Different Environmental Samples. BiosciBiotechnol Res Asia. 2023;20(4):1303-15.
CrossRef - Etienne D, Shareck, F, Hurtubise Y, Daneault C, Beauregard. M. Homologous cloning, expression and characterization of a laccase from Streptomyces coelicolor and enzymatic decolourisation of an indigo dye. ApplMicrobiolBiotechnol. 2008;79: 597-603.
CrossRef - Babadi ZK, Narmani A, Ebrahimipour GH, Wink J. Combination and improvement of conventional DNA extraction methods in Actinobacteria to obtain high-quantity and high-quality DNA. Iran J Microbiol. 2022; 14(2):186-193.
CrossRef - Cornell CR, Marasini D, Fakhr MK. Molecular Characterization of Plasmids Harbored by Actinomycetes Isolated From the Great Salt Plains of Oklahoma Using PFGE and Next Generation Whole Genome Sequencing. Front Microbiol. 2018; 9:2282.
CrossRef - Annette K, Korner H, Kneist S, Eschrich K. Oral actinomycetes isolates forming red colonies on brain heart blood agar can be unambiguously classified as A. odontolyticus by macroscopic examination. J Clin Microbiol.2003; 41(8): 3729-3731.
CrossRef - Vantamuri AB, Kaliwal BB. Decolourization and biodegradation of Navy blue HER (Reactive Blue 171) dye from Marasmius sp. 3 Biotech. 2017;7(1):48
CrossRef - de Almeida AP, Macrae A, Ribeiro BD, do Nascimento RP. Decolorization and detoxification of different azo dyes by Phanerochaetechrysosporium ME-446 under submerged fermentation. Braz J Microbiol. 2021;52(2):727-738.
CrossRef - Durão P, Gülereşi D, Proença J, Gordo I. Enhanced Survival of Rifampin- and Streptomycin-Resistant Escherichia coli Inside Macrophages. Antimicrob Agents Chemother. 2016;60(7):4324-32.
CrossRef - Carneiro-da-Cunha MD, Lima Filho JL, Campos-Takaki GM. Protoplast formation and regeneration from Streptomyces clavuligerus NRRL 3585 and clavulanic acid production. Braz J Microbiol. 2002;33:347-51.
CrossRef - Sandgrind S, Li X, Ivarson E, Ahlman A, Zhu LH. Establishment of an Efficient Protoplast Regeneration and Transfection Protocol for Field Cress (Lepidiumcampestre). Front Genome Ed. 2021;3:757540
CrossRef - Guo H, Zheng B, Jiang D, Qin W. Overexpression of a Laccase with Dye Decolorization Activity from Bacillus sp. Induced in Escherichia coli. J MolMicrobiolBiotechnol. 2017; 27(4):217-227.
CrossRef - Liu Y, Huang L, Guo W, Jia L, Fu Y, Gui S, Lu F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochem. 2017; 53:125-34.
CrossRef - Kaur R, Salwan R, Sharma V. Structural properties, genomic distribution of laccases from Streptomyces and their potential applications. Process Biochem. 2022; 118:133-44.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.



