

How to Cite | Publication History | PlumX Article Matrix
Chubanaro Aier and Jeyaparvathi Somasundaram*
and Jeyaparvathi Somasundaram*
Department of Zoology, St. Joseph University , Chumoukedima, Nagaland, India.
Corresponding Author E-mail:jeya5001@gmail.com
ABSTRACT: The study examined the midgut microbial composition of Eri silkworms, or Samia ricini, in order to clarify the richness and variety of their bacterial community throughout several gut compartments and developmental phases. A wide variety of microbial taxa that are essential for immunity, nutritional absorption, and digestion were found by us using both culture-dependent and culture-independent approaches. With 22% of the microbial population, Enterococcus took the lead, followed by Acinetobacter (10%), Escherichia-Shigella (9%), and Proteiniphilum (9%). The intricacy of the microbiota was further demonstrated by uncultured bacterial species (9%) and chloroplast-associated bacteria (12%). The midgut was shown to have the largest bacterial load and diversity, making it the main microbial hotspot. A rich and intricate bacterial ecosystem was confirmed by analysing the microbial diversity using Shannon–Wiener and Simpson diversity indices by 16S rRNA gene sequencing. The ecological importance of these microbial communities, their function in host metabolism, and the possibility of using probiotics to improve silk output in sericulture were also investigated in this work. The results shed more light on the gut microbiome of lepidopterans and its wider effects on nutrition, insect health, and sustainable sericulture methods.
KEYWORDS: Acinetobacter; Enterococcus; microbial diversity; Proteiniphilum; Samia ricini; gut microbiota; 16S rRNA sequencing
| Copy the following to cite this article: Aier C, Somasundaram J. Microbial Diversity in the Midgut of the Eri Silkworm (Samia ricini): Ecological Significance and Implications for Sericulture. Biotech Res Asia 2025;22(1). |
| Copy the following to cite this URL: Aier C, Somasundaram J. Microbial Diversity in the Midgut of the Eri Silkworm (Samia ricini): Ecological Significance and Implications for Sericulture. Biotech Res Asia 2025;22(1). Available from: https://bit.ly/4kD8ieV |
Introduction
The Eri silkworm, also known as Samia ricinii, is an important ecological value in addition to being a major contributor to the silk industry. This insect is native to Asia, especially in China and India. It is used in sericulture to produce Eri silk, which is prized for its special qualities, such as its durability and warmth. In addition to its commercial worth, the Eri silkworm is an important component of regional ecosystems because it participates in complex food webs.By providing food for different predators, such as birds and other insects, it promotes biodiversity. By controlling herbivory, it also promotes healthier flora by influencing the growth of host plants, particularly the castor bean plant (Ricinus communis). This highlights the interdependence of biological systems by making the Eri silkworm more than just a commercial commodity but also an essential part of its ecological habitats.In insects, gut symbiotic microbes perform a number of physiological functions related to host metabolism, such as food digestion, the provision of vital nutrients that are lacking in food, the inhibition of pathogen invasion by boosting the host immune system, the breakdown of pesticides and phytotoxins, the synthesis of antibiotics, and competition for scarce nutrients.1 In order to further understand the intricate relationships between host organisms and their symbiotic microbiota, it is essential to comprehend the microbial communities that inhabit the midgut of the Eri silkworm (S.ricinii). Instead of being passive inhabitants, these microbial populations are essential to the host’s metabolism, digestion of nutrients, and general health. The pH levels, oxygen availability, and substrate types of the midgut provide a special habitat that influences the makeup and activity of these microbial groups.Studies on insect gut bacteria have utilized both cultivable and culture-independent approaches.2 Recent developments in molecular methods, including microbial sequencing and metagenomics, have shown how diverse the bacteria and other microbes that live in this gut niche are. In order to clarify the ecological relevance of the microbial communities in the midgut of Eri silkworms and their possible ramifications for sericulture methods, this study attempts to present a thorough investigation of these populations. A comparatively objective perspective on the composition of intricate microbial communities can be obtained by culture-independent methods like 16S metagenomics analysis.
Numerous studies have lately reported the presence of complex and numerous microbial communities in the guts of lepidopteran insects using high-throughput sequencing techniques.3,4 To improve our knowledge of symbiosis in insect hosts, this study aims to shed light on the complex link between the Eri silkworm and its gut bacteria.
Materials and Methods
The Eri Silkworm eggs were collected from Department of Sericulture, Dhansiripar,Dimapur, Nagaland. The eggs were incubated in sterile petri dishes at the silkworm rearing room under Department of Zoology at St. Joseph University, under a light and dark cycle of 16:8 hours, with a rearing temperature of 78.8°F and 70% relative humidity, until they hatched. Following the conventional rearing protocol,5 the first instar larvae were hatched and then placed into rearing boxes that had been formalin-sterilized. They were then kept on castor leaves. Tender leaves were fed to the larvae in their first instar till their second and third instars. Mature castor leaves were fed to the larvae in their fourth and fifth instars.Using the third, fourth, and fifth larval and egg stages, the gut flora of S. ricini was isolated and removed. For each stage, five healthy larvae were selected and starved for 24 hours prior to the intestines being removed. After rinsing the larvae in double-distilled water for 30 seconds, 70% (v/v) ethanol for 60 seconds, and then double-distilled water for another 30 seconds, the disinfectant was removed.Using sterile micro scissors in a laminar flow exposed to UV light, the sterile larvae were sliced open to remove the stomach.A sterile homogeniser was used to homogenise the midgut in a single 1.5 ml Eppendorf tube. For a short time, the homogenised midgut was centrifuged to remove any remaining food particles. The supernatant was then used in repeated dilutions and as an inoculum for bacterial enrichment when necessary.To separate gut bacteria, the following culture media were acquired from HiMedia Laboratories Pvt. Ltd., Mumbai, India: The nutrient agar (NA) medium consisted of 20 g of agar, 3 g of beef extract, 5 g of peptone, 5 g of NaCl, and 1,000 ml of distilled water.The media was autoclaved at 121°C for twenty minutes followed by serial dilution. The serially diluted gut homogenates from the various compartments were used to create three sets of inoculations on agar plates.
The contaminated plates were incubated at 37°C for 24 to 48 hours. To obtain the desired level of purity for each culture, a single representative isolate of each morphotype was streaked over corresponding agar plates several times. The size, colour, and form of the bacterial colonies were used to identify them.After a 24-hour fast, fifth-instar S. ricini larvae were pre-chilled for one to two minutes to immobilise and make handling easier. The insects were thoroughly sterilised in 70% ethanol for 60 seconds, followed by a rinse in double-distilled water before dissection. Each insect was promptly dissected as described above, put into a 1.5 ml sterile Eppendorf tube with a sterile 0.85% NaCl solution, covered with parafilm, and kept at -20°C until it was needed again.To identify the bacteria, separate representative pure colonies were chosen based on 16S rRNA gene probes. A 24-hour culture of individual pure bacterial isolates was conducted in nutritive broth at 37°C. To separate the pellet and supernatant, the broth cultures were centrifuged at 13,000 rpm following a 24-hour growth period. DNA was extracted using a technique appropriate for the kind of sample using commercially available kits like QIAGEN, ZYMO RESEARCH, and ThermoFisher. Primers16sF:-(5’AGAGTTTGATGMTGGCTCAG3′)and16sR:- (5’TTACCGCGGCMGCSGGCAC3′) were used to amplify the 16srRNA gene. For amplification, 40 ng of extracted DNA and 10 pM of each primer were utilised. The following is how the PCR was carried out: Initial Denaturation: 95°C, 25 cycles under the specifications listed below: 15 seconds of denaturation at 95°C, 15 seconds of annealing at 60°C, and 2 minutes of elongation at 72°C.Ten minutes of final extension at 72°C and holding at 4°C.To determine the diversity and distribution of gut microbes across the midgut, the identified gut bacteria isolated from the aforementioned section were examined using the Shannon–Wiener index (H’). The data on the gut bacteria of eri silkworms that were extracted from midgut sections over the course of their life cycle were subjected to diversity analysis. The Shannon–Wiener and Simpson diversity indices were calculated using the procedures described.6
Results
Microbial Diversity Across Developmental Stages and Gut Compartments
A very complex and varied bacterial community was found in the study of the midgut microbial composition of Samia ricini (Eri silkworm), with notable differences seen between various gut compartments and developmental phases. We gained a thorough grasp of the gut microbiota by utilising both culture-dependent and culture-independent approaches, emphasising the dynamic character of bacterial colonisation in S. ricini.
Dominant Microbial Taxa
The bacterial community structure was analyzed, and Enterococcus emerged as the predominant genus, comprising 22% of the total microbial population. This was followed by Acinetobacter (10%), Escherichia-Shigella (9%), and Proteiniphilum (9%). The substantial representation of these bacterial genera suggests their potential functional roles in digestion, nutrient assimilation, and host metabolism.
The study found a noteworthy percentage of chloroplast-associated bacteria (12%) in addition to these dominant species, which might have resulted from the host’s plant-based diet. The intricacy of the gut microbiota is further highlighted by the presence of uncultured bacterial species (9%) and the necessity of using sophisticated genomic techniques to describe these unknown microbial communities.
Gut Compartment-Specific Variability
The midgut was shown to be the main microbial hotspot, however the microbial composition varied greatly between the various gut compartments. Because of its vital role in digestion and nutrition absorption, this region showed the highest bacterial diversity and load. The midgut’s crucial role in preserving a stable and functionally active microbial ecology was highlighted by the relatively reduced microbial populations in other gut compartments, such as the foregut and hindgut.
Influence of Developmental Stages on Microbial Composition
The study also showed that S. ricini’s gut microbiota varies in composition depending on its developmental stage. Bacterial diversity was higher in larval stages than in later stages, indicating that microbial colonisation is more active in the early phases of development. The existence of particular bacterial taxa at particular phases suggests possible symbiotic relationships that could support digestion, growth, and immunity.
Microbial Diversity Analysis and Statistical Validation
16S rRNA gene sequencing was used to measure microbial diversity, and diversity indices including the Simpson and Shannon-Wiener indices verified a diverse and rich bacterial environment. The notion that the midgut is the main centre of microbial activity was further confirmed by the high diversity index values found there. The heterogeneity in bacterial composition was further supported by Principal Coordinate Analysis (PCoA), which showed clear clustering of microbial communities according to gut compartment and developmental stage.
Potential Functional Roles of Identified Bacteria
Since members of the genus Enterococcus are known to produce bacteriocins, which inhibit harmful bacteria, their prevalence suggests that they play a role in both antimicrobial defence and carbohydrate metabolism. While Escherichia-Shigella species are probably involved in fermentation and other digestive processes, Acinetobacter has been linked to nitrogen metabolism. Proteiniphilum’s presence would suggest that it plays a part in protein breakdown, which helps S. ricini use nutrients effectively. Furthermore, the discovery of bacteria linked to chloroplasts raises the possibility of interactions between the gut microbiota and plant-based foods consumed by silkworms.
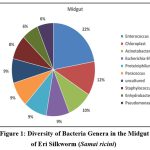 |
Figure 1: Diversity of Bacteria Genera in the Midgut of Eri Silkworm (Samai ricini) |
Discussion
The gut microbiota of the Samia ricini, or Eri silkworm, is crucial to its development, health, and overall health. This complex association of microbes not only facilitates digestion but also maintains the immunity of the insect, demonstrating an incredible symbiosis between the host and its microbial residents.Since these silkworms have a diet composed predominantly of plant material, their gut becomes an active ecosystem where helpful bacteria, including Enterococcus and Acinetobacter, thrive. These bacteria break down the complex nutrients into forms that are easier to absorb, as well as serve as a protective mechanism against pathogenic infection. This mutualism underscores the significance of diet in structuring the gut microbiome, since the composition of the plants eaten has direct implications for the diversity and function of the microbial community. In addition, the dynamic nature of the gut microbiota across the silkworm life cycles indicates its plasticity and resilience. As the Eri silkworm ages, the gut microbiome also undergoes changes as it responds to environmental exposure and dietary shifts. This flexibility not only increases the insect’s potential to survive but also presents new avenues for enhancing sericulture practice. Through the use of probiotics and insight into the complex interactions within the gut microbiome, we can create sustainable means of enhancing silkworm health and productivity, ultimately contributing to the silk industry. Essentially, the gut microbiota of Samia ricini is not merely an assemblage of bacteria but a vital companion in the life of the insect, determining its health, growth, and capacity to adapt to its surroundings. This knowledge can pave the way for novel solutions in sericulture, ensuring both the health of the silkworms and the sustainability of silk production. The gut microbiota of Samia ricini, or the Eri silkworm, is vital for its development, well-being, and general well-being. The intricate population of microbes has a major function in digestion and enhances the immune system of the insect, providing an interesting example of symbiosis between the host and its microbial inhabitants. Since Eri silkworms have a diet rich in plant material, the gut is converted into a colorful ecosystem with beneficial microbes such as Enterococcus and Acinetobacter. These microbes help to decompose complex nutrients so that they are easily absorbed by the body while, at the same time, acting as a safeguard against unwanted pathogens. This mutualistic process highlights the importance of diet in influencing the gut microbiome since the type of plants eaten directly influences the diversity and functionality of the microbial community.
Furthermore, the dynamic nature of the gut microbiota across the life cycle of the silkworm reflects the adaptability and robustness of its microbiome. As the Eri silkworm goes through its developmental stages, its gut microbiome changes due to variations in diet and environmental stimuli. This flexibility not only improves the survival of the insect but also offers thrilling possibilities for the development of sericulture practices. Through the utilization of the advantages of probiotics and the implications of the complex interactions of the gut microbiome, we are able to create sustainable approaches for enhancing silkworm health and productivity, thereby deriving benefits for the silk industry.
In short, the gut microbiota of Samia ricini is not merely an assembly of bacteria; it is a crucial partner in the insect’s life process, determining its health, growth, and resistance to environmental shifts. This insight opens the door to new avenues in sericulture, promoting the health of the silkworms as well as the sustainability of silk farming.
Conclusion
The midgut microbial composition of S. ricini (Eri silkworm) throughout several gut compartments and developmental stages is thoroughly examined in this study. We discovered a varied bacterial community that was dominated by Enterococcus, Acinetobacter, Escherichia–Shigella, and Proteiniphilum using both culture-dependent and culture-independent methods. The intricacy of the gut microbiota is further highlighted by the existence of uncultured species and bacteria linked to chloroplasts. With the largest bacterial diversity and load, the midgut has been identified as the main microbial hotspot. Bacteria are crucial for digestion, nutrient absorption, and host metabolism.The ecological significance of the gut microbiota in S. ricini is highlighted by our findings, which also imply that some bacterial groups may be useful probiotics to enhance silk production in sericulture. This work provides important new information about the dynamic interactions between microbial and host communities by utilising diversity indices and 16S rRNA gene sequencing. The functional characteristics of these microorganisms and their possible uses in boosting S. ricini growth, immunity, and silk yield should be investigated in future research. Utilising and comprehending insects’ gut microbiome can also lead to effective and sustainable sericulture methods.
Acknowledgement
The author would like to thank St. Joseph University for providing the Lifesciences Lab for carrying out the research work and Dr. S. Jeyaparvathi for guiding throughout the work process.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
Chubanaro Aier – Sample collection, carrying out research work.
S Jeyaparvathi – Research Guide, farmed working methodology and contributed in carrying out the research work efficiently.
References
- Arora AK, Douglas AE. Hype or opportunity? Using microbial symbionts in novel strategies for insect pest control. Journal of Insect Physiology. 2017; 103:10-17.
CrossRef - Chen B, Yu T, Xie S, Comparative shotgun metagenomic data of the silkworm Bombyx mori gut microbiome. Scientific Data. 2018;5(1).
CrossRef - Chen, B., Teh, B. S., Sun, C., Hu, S., Lu, X., Boland, W., & Shao, Y. (2021). Biodiversity and activity of the gut microbiota across the life history of the insect host. Nature Communications, 12, 2788.
- Dubey, A. K., & Ben-Yosef, M. (2020). Enterococcus as a beneficial symbiont in insect gut microbiota: Implications for host health and disease management. Microbial Ecology, 79, 883–899.
- Gandotra S, Kumar A, Naga K Bacterial community structure and diversity in the gut of the muga silkworm, Antheraea assamensis (Lepidoptera: Saturniidae), from India. Insect Molecular Biology. 2018;27(5):603-619
CrossRef - Hammer, T. J., Janzen, D. H., Hallwachs, W., Jaffe, S. P., & Fierer, N. (2017). Caterpillars lack a resident gut microbiome. PNAS, 114(38), 9641-9646.
CrossRef - Indiragandhi, P., Anandham, R., Madhaiyan, M., Poonguzhalan, R., Kim, G. H., Saravanan, V. S., & Sa, T. M. (2007). Diversity of culturable bacteria associated with the gut contents of larvae and adults of rice swarming caterpillar. Journal of Basic Microbiology, 47, 439-447.
- Meth T, Gogoi H. Rearing of eri silkworm (Samia cynthia ricini) (Lepidoptera: Saturniidae) in arunachal pradesh: a study in papumpare district. Bioresources. 2016; 3:46–52.
- Montagna, M., Chouaia, B., Mazza, G., Prosdocimi, E. M., Crotti, E., Mereghetti, V., et al. (2016). The microbiota associated with Antheraea polyphemus larvae. Microbiome, 4, 38.
- Pinto-Tomás AA, Sittenfeld A, Uribe-Lorío L Comparison of Midgut Bacterial Diversity in Tropical Caterpillars (Lepidoptera: Saturniidae) Fed on Different Diets. Environmental Entomology. 2011;40(5):1111-1122.
CrossRef - Sevim E, Çocar M, Sezgin FM, Sevim A. Aerobic gut bacterial flora of Cydia pomonella (L.) (Lepidoptera: Tortricidae) and their virulence to the host. Egyptian Journal of Biological Pest Control. 2018;28(1).
CrossRef - Swathiga G, Umapathy G, Parthiban KT, Angappan K. Growth response of different eco races of ERI silkworm reared on various castor genotypes. Entomol. Zool. J. 2019; 7:1406–1410.
- Zelles L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biology and Fertility of Soils. 1999;29(2):111-129.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.



