

Manuscript accepted on : February 25, 2009
Published online on: 22-06-2009
Cytogenetic Studies on Fluted Pumpkin (Telfairia Occidentalis Hook. F.)
J. C. Onovo and M. I. Uguru
Department of Crop Science, University of Nigeria, Nsukka Nigeria.
Corresponding Author E-mail: onovojos@yahoo.com
ABSTRACT: The study was initiated to determine the cytological status of fluted pumpkin (Telfairia occidentalis). The analyses were carried out in the mitotic metaphase stage using the cold-induced pretreatment method. The results revealed diploid chromosome number of 2n = 22. However, a diploid chromosome number of 2n = 22 + 1 was observed in about 4% of the cells studied. The chromosome morphology was described on the basis of the centromere position. The ratio of the sum of the short arm length to the total chromosome length (TF%) of 36.9% was obtained. The centromic index (F%) of 1.52 and 45.19 were recorded. The karyotype consisted of 5 metacentric, 4 submetacentric, 1 subtelocentric and 1 telocentric chromosomes. With the exception of the 10th and 11th chromosomes that were subtelocentric and telocentric, respectively, the chromosomes were mostly metacentric and submetacentric. The frequency of the diploid chromosome number of 23 was low, and as such could be regarded as one of those occasional abnormalities that occur in plant species. This could be an aneuploid that was produced accidentally due to an extra chromosome (2n + 1).
KEYWORDS:
Cytogenetics; chromosome; fluted pumpkin; Telfairia occidentalis; karyotype
Download this article as:| Copy the following to cite this article: Onovo J. C, Uguru M. I. Cytogenetic Studies on Fluted Pumpkin (Telfairia Occidentalis Hook. F.). Biosci Biotechnol Res Asia 2009;6(1) |
| Copy the following to cite this URL: Onovo J. C, Uguru M. I. Cytogenetic Studies on Fluted Pumpkin (Telfairia Occidentalis Hook. F.). Biosci Biotechnol Res Asia 2009;6(1). Available from: https://www.biotech-asia.org/?p=7812 |
Introduction
The fluted pumpkin (Telfairia occidentalis Hook. f.) belongs to the family cucurbitaceae which consists of 90 genera and 750 species (Purseglove, 1984). It is commonly grown as an annual vegetable but sometimes perennial ratoon shoots do exist. It is mainly used as a leaf vegetable in Nigeria. The seeds contain over 40% oil, making it a potential raw material for chemical and pharmaceutical industries (Okoli and Mgbeogwu, 1983; Asiegbu, 1987).
Karyotype is a reliable taxonomic tool for characterizing plant and animal species. It has served as a genetic marker in some crops for elucidating plants’ systematics and evolution (Robert and Foster, 1971; Doudrick et al., 1995). For instance, information on the taxonomic relationship of Araucaria auraucana and A. angustifolia was obtained by Cardemil et al., (1984) from their karyotypes.
Falistocco et al., (1995) with the aid of karyotypes, characterized the chromosomes of Medicago and provided further evidence of the autotetraploid origin of the species. Karyotype is used for reference when screening and describing chromosome mutations, and for studying the relationship between different species (Dyer, 1979).
The cytological features of fluted pumpkin, which are urgently needed for its gene mapping and characterization of the morphotypes are yet to be comprehensively studied. There is a report in the literature (Okoli and Mgbeogwu, 1983) that fluted pumpkin has a chromosome number of 2n = 22, but the report did not contain karyotypic details of the crop. The present study extends beyond numerical count to karyotyping and detailed morphology of the chromosomes. A detailed knowledge of the karyotype of the fluted pumpkin will facilitate the characterization of the different morphotypes that exist in the species. The present study was initiated to determine the cytological status of fluted pumpkin and as a corollary, to establish how such cytologenic information would be utilized in the improvement of the crop.
Materials and Methods
The study was initiated in May, 2005 in the Laboratory of the Department of Crop Science, Faculty of Agriculture, University of Nigeria, Nsukka and completed in November, 2007 at the Cytogenetic Laboratory of the Department of Genetics, Faculty of Agriculture, Minia University, Egypt. Mitotic analysis of Telfairia occidentalis was carried out using the cold-induced pretreatment method (cold treatment) as an integral part of the conventional techniques. Tender root tips of germinating seeds extracted from fluted pumpkin pods were harvested (excised) at 11.00 am when mitotic activities are believed to be high (Jackson, 1962), using sterilized forceps and scalpel. The root tips, nearly 1cm in length were washed with distilled water and cooled at 4oC for 48 hours in a refrigerator.
The root tips were fixed with freshly prepared farmer solution (3 parts ethyl alcohol : 1 part glacial acetic acid) for 24 hours and stored in 70% ethanol. They were rinsed with distilled water twice and hydrolysed in 18% HCl for 10 minutes at 60oC and rinsed in distilled water. The meristematic region of a young root tip was squashed in a drop of aceto-carmine (1%) on a slide. The cover slip was tapped gently with the end of a dissecting needle to spread the cells. The cells were then flattened by supplying firm pressure with the thumb on the slide that was placed between folds of filter paper. The slides were examined under a compound microscope. The slides that had well spread metaphase were photographed at x1000 using an Olympus, BX 51 model of photomicroscope equipped with Olympus C 4040 digital camera. Faluyi (1992) observed that photomicrographic documentation is very useful for proper identification.
Measurements of the short arm (s), long arm (l) and the chromosome length were made for each chromosome. The arm ratio (r = l/s) was calculated and used to classify the chromosomes as proposed by Leven et al., (1961). The karyotype was constructed by arranging the chromosomes in order of decreasing length. The chromosomes were also organized into metacentric, submetacentric, subtelocentric and telocentric according to their arm ratios. The chromosomes were measured in microns and also classified into large, medium or small on the basis of their length (Fukui and Nakayama, 1996) as follows:
Type Chromosome length
Large (L – type) 8.0 – 10.0 µm
Medium (M – type) 3.1 – 7.9 µm
Small (S – type) 2.0 – 3.0 µm
The centromeric index (F%) was calculated as the ratio of short arm length to the total chromosome length expressed in percent (Huziwara, 1962). The sum of short arm length / total chromosome length relation (TF%) was also calculated.
Results
Mitotic chromosome count of 2n = 22 was recorded (Plate 1). The chromosomes showed much variability in size ranging from 3.30 and 7.48 µm (x = 5.04 µm) in length but less variable in structure. The photomicrographs showed clear evidence that the fluted pumpkin is a diploid. However, about 4% of the 123 observed cells showed mitotic chromosome number of 2n = 22 + 1 (Plate 2).
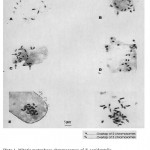 |
Plate 1: Mitotic metaphase chromosomes of T. occidentalisshowing diploid chromosomes (2n=22) in six cells A, B, C, D, E and F.
|
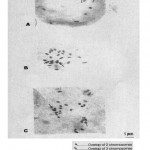 |
Plate 2: Mitotic metaphase chromosomes of T. occidentalisshowing aneuploidy (2n=22+1) in three cells A, B and C.
|
Table 1 and Plate 3 show the karyotype and the total chromosome length of the eleven pairs of the chromosomes of fluted pumpkin. Chromosome 1 was the longest with a lightly stained terminal region and it is metacentric. The chromosome 2 has lightly stained proximal regions on both the long and short arms, and is also metacentric. Chromosomes 3 and 4 are both metacentric with distally located heterochromatic segments. Chromosome 5 was submetacentric and had a nucleolus organizer region (NOR) close to its distal region. It has a corresponding homologue (homomorphic with respect to length) that lacked a NOR and its distal end darkly stained. It can also be distinguished by having the largest size in the submetacentric group. Chromosome 6 has its distal end darkly stained and is submetacentric. Chromosome 7 has the smallest size in the metacentric group. Chromosomes 8 and 9 are submetacentric. Chromosome 10 is subtelocentric while chromosome 11 is telocentric. When the total chromatin length was considered, the minimum of 3.30 and maximum of 7.48 µm were recorded. The F% ranged from 1.52 to 45.19%, and a TF% of 36.9% was obtained.
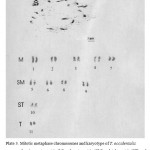 |
Plate 3: Mitotic metaphase chromosomes and karyotype of T. occidentalis showing metacentric (M), submetacentric (SM), subtelocentric (ST) and telocentric chromosomes. |
Table 1: Measurement of 11 pairs of mitotic chromosomes of T. occidentalis in respect of chromosome length, arm ratio, F% and centromere position.
| Chromosome Total
pair number chromosome Long arm length (µm) |
Short arm (µm) |
Arm ratio |
F%
|
Centromere position |
|
| 1 7.48 + 0.29
2 6.60 + 0.25 3 6.15 + 0.18 4 5.85 + 0.20 5 5.25 + 0.12 6 4.85 + 0.11 7 4.50 +0.16 8 4.16 + 0.18 9 3.75 + 0.10 10 3.55 + 0.11 11 3.30 + 0.11 Total 55.44 Mean 5.04
|
4.10 + 0.19
3.73 + 0.15 3.63 + 0.12 3.55 + 0.15 3.25 + 0.11 3.15 + 0.09 2.40 + 0.15 2.78 + 0.20 2.50 + 0.18 2.65+ 0.16 3.25 + 0.10 34.99 3.18
|
3.38 + 0.17
2.87 + 0.16 2.52 + 0.15 2.30 + 0.11 2.00 + 0.07 1.70 + 0.16 2.10 + 0.14 1.38 + 0.18 1.25 + 0.17 0.90 + 0.40 0.05 + 0.02 20.45 1.86
|
1.21
1.30 1.44 1.54 1.63 1.85 1.14 2.01 2.00 2.94 65.0
|
45.19
43.48 40.98 39.32 31.05 35.05 46.67 33.17 33.33 25.35 1.52
|
M
M M M SM SM M SM SM ST T
|
Discussion
Diploid chromosome number of 2n = 22 is in agreement with an earlier report (Okoli and Mgbeogwu, 1983). The existence of aneuploids was however not reported by the authors. They did not also have any report on the karyotypic details of fluted pumpkin. The karyotype constructed from the study had the formula, 2n = 22 = 5m + 4sm + 1st + 1t, which implies that the karyotype of the species consists mainly of metacentric and submetacentric chromosomes, indicating little structural variability among the chromosomes. Olorode et al. (1982) reported that the position of the centromere is often useful in distinguishing one chromosome (or a homologous pair) from another.
Based on the classification of Fukui and Nakayama (1996), the diploid genotype of fluted pumpkin would be characterized as the medium type (M – type) chromosomes as they range from 3.30 – 7.48 µm. The medium chromosome length would appear to suggest that advancement of this taxon has taken place in cultivation. The wide F% range of 1.52 to 45.19 in addition to the presence of metacentric, submetacentric and telocentric chromosomes provide additional support to this proposition.
The telocentric position of the centromere of the chromosome 11 may have been brought about by deletion and consequent reduction in the size of one of the arms of the chromosome, causing shifting of the centromere position and reduction in the absolute size of the chromosome (Sinha and Roy, 1979). The presence of a telocentric chromosome pair in addition to median and submedian chromosomes depict asymmetrical karyotypes in the diploid fluted pumpkin. This further validates that fluted pumpkin has gone beyond the primitive stage to some level of advancement over the years. Germplasm collection of this useful crop will be worthwhile at this stage of development.
Aknowledgements
The authors which to thank Prof. Abdel Tawab M. Ata of the Deparment of Genetics, Faculty of Agriculture, Minia University, Egypt for his assistance in the preparation of slides, statistical analysis and for availing us his expertise in photomicrography. We are grateful to the Deparment of Genetics, Faculty of Agriculture, Minia University, Egypt for permitting us to carry out part of the research work in the Cytogenetic Laboratory of the Depatment.
References
- Asiegbu, J.E. (1987). Some Biochemical Evaluation of Fluted Pumpkin Seeds. Sci. Food Agric. 40: 152-155
- Cardemil, L.; Salas, E. and Godoy, M. (1984). Comparative Study of the Karyotype of South American Species of Araucaria. The Journal of Heredity 75: 121-125
- Doudrick, R.L., Heslop-Harrison, J.S., Nelson, C.D., Schmidt, T.,Nance, W.L. and Schwarzcher, T. (1995). Karyotype of Slash Pine (Pinus elliotii var. elliotii). Using Patterns of Florescence In situ
- Hybridization and Fluorochrome Banding. J. Hered. 86: 289-296.
- Dyer, A.F.(1979). Investigating Chromosomes. London, Edward Arnold. Falistocco, E.; and Falcinelli, M. and Veronesi, F. (1995). Karyotype and C-
- banding pattern of mitotic chromosomes in alfalfa, Medicago sativa L. Plant Breeding 114:451-453
- Faluyi, J.O. 1992. Principles and Practice of Photomicrography. Anchorprint Ltd. Ibadan, 102 pp
- Fakui, K. and Nakayama, S. 1996. Plant Chromosomes: Laboratory Methods. CRC Press. 298pp.
- Huziwara, Y. (1962). Karyotype Analysis in Some Genera of Compositae viii. Further Studies on the Chromosomes. Asher: Amer. J. Bot. 49: 116-119.
- Jackson, R.C. (1962). Interspecific Hybridization in Haplopappus and its Bearing on Chromosomes Evolution in the Blepharodon Section. American Journal of Botany 49 (2): 119-132.
- Levan, A.K., Fredgar and A.A. Sandberg (1961). Nomenclature for Centromic Position on Chromosomes. Hereditas 52: 201- 220.
- Okoli, B.E. and Mgbeogwu, C.M. (1983): Fluted Pumpkin Telfairia occidentalis, West African Vegetable Crop. Economic Botany 3(2): 145-149.
- Olorode, O., Fatunla, and Adegoke (1982). Introductory Genetics. University of Ife Press Ltd, 209pp.
- Puresglove, J.W. (1984). Tropical Crops. Dicotyledons. Volumes 1 and 2. English Language Book Society and Longmans, London. p 100.
- Robert, E. and Foster, I.I. (1971). Downy Midew Immunity as a Genetic Marker for Lettuce Breeding. J. Hered. 62: 316-318.
- Sinha, S. S. N. and Roy, H. 1979. Cytological Studies in the Genus Phaseolus. Mitotic analysis in the fourteen species. Cytologi 44: 191-199.

This work is licensed under a Creative Commons Attribution 4.0 International License.



