

How to Cite | Publication History | PlumX Article Matrix
Chlorophyll Fluorescence of the Desert Plant Calotropis Procera Grown Under Water Deficit Stress
Abdellah Akhkha
Department of Biology, Faculty of Science, Taibah University, Al-Madinah Al-Munawwarah, Kingdom of Saudi Arabia.
Corresponding Author E-mail: abdellah99@gmail.com
ABSTRACT: The effectiveness of the photosynthetic machinery of the desert plant Calotropis procera grown under different water deficit regimes was investigated by chlorophyll fluorescence technique in order to understand how such desert plant withstand water limitations without a irreversible damage to the integrity of the Photosystem II of the photosynthetic apparatus. At the onset of stress (first harvest), all chlorophyll fluorescence parameters Fo, Fm, Fv / Fm and Fv / Fo, were reduced in both low and mild water deficit regimes 30% and 50% respectively. However, in the second harvest, stressed leaves showed some recovery and maximal fluorescence Fm was increased at the low water regime. All chlorophyll fluorescence ratios were also increased in both low and mild water deficit regimes. At the third harvest, Fo and Fm were still higher in low water regime, but decreased or reached a level similar to that of well-watered plants respectively at mild water regime. Chlorophyll fluorescence ratios at low water regime were similar to that of the well-watered plants but higher at mild water regime. Generally, the photosynthetic apparatus was reversibly affected by both limited water regimes on the onset of water stress; however, stressed leaves were recovered and the photosynthetic machinery became sometimes similar to that of well-watered plants or even more efficient at other times. Chlorophyll Index was increased in all water regimes and at all harvests. The relationships between Chlorophyll Index and the different chlorophyll fluorescence parameters are discussed.
KEYWORDS: Chlorophyll fluorescence; Calotropis procera; water deficit regimes
Download this article as:| Copy the following to cite this article: Akhkha. A. Chlorophyll Fluorescence of the Desert Plant Calotropis Procera Grown Under Water Deficit Stress. Biosci Biotechnol Res Asia 2009;6(2) |
| Copy the following to cite this URL: Akhkha. A. Chlorophyll Fluorescence of the Desert Plant Calotropis Procera Grown Under Water Deficit Stress. Biosci Biotechnol Res Asia 2009;6(2). Available from: https://www.biotech-asia.org/?p=8752. |
Introduction
When green plants are subject to water deficit, many physiological processes are perturbed such as photosynthesis, respiration, translocation, partitioning of metabolites, stomatal behaviour, protein synthesis, cell division, cell elongation and cell wall synthesis. Therefore, plants under water deficit stress have a limited growth and low productivity (Ingram & Bartels, 1996). Such effects are well investigated in crop plants although understanding the mechanisms involved is still limited.
In the contrary to crop plants, desert plants are less studied and more work is still to be carried out to investigate how desert plants survive water deficit stress and limit the effect of the stress on growth and productivity. Many techniques are used to sense the response of desert plants to water deficit stress such as photosynthesis rate, chlorophyll content, Chlorophyll fluorescence and respiration. (Akhkha, 2009, in press). Photosynthesis rate and chlorophyll content were found to be reduced in many crop plants and some desert plants (HongBo et al., 2005; Hamerlynck et al., 2000). However, some authors reported that chlorophyll fluorescence is a more reliable technique to sense environmental stress including water deficit stress (Rong-hua et al., 2006; Araus et al., 1998; Araus & Hogan, 1994).
Many authors investigated the effect of water stress on chlorophyll fluorescence as an indicator of the effect of stress on the photosynthetic machinery (Massacci, et al., 2008; Souza et al., 2004; Vassilev & Manolov, 1999). Chlorophyll fluorescence was proven to be a powerful method for measuring the effect of water stress on a number of photosynthetic traits such as initial fluorescence Fo, maximum fluorescence Fm, maximum primary yield of photochemistry of photosystem II (Fv / Fo), maximum quantum yield of photosystem II (Fv / Fm) (Reference).
Water stress was found to affect chlorophyll fluorescence in many crop plants. Rong-hua et al. (2006) reported that drought reduced all chlorophyll fluorescence parameters in four barley genotypes indicating damage to the photosynthetic apparatus. Yang et al. (2006) also observed a decrease in chlorophyll fluorescence parameters in many wheat genotypes. However, the level of decrease was not similar in all barley and wheat genotypes, indicating the presence of water deficit tolerance in some genotypes. Similarly, all chlorophyll fluorescence parameters were reduced in maize plants subjected to water deficit stress (Haveau & Lannoye, 1983).
In contrast, the scientific literature showed that very few desert plants were used to examine the reliability of this method to study the effect of water deficit stress on desert plant species. For example, chlorophyll fluorescence represented by the photochemical efficiency of photosystem II (Fv / Fm) in the desert plant Larrea tridentate was found to be reduced in response to water deficit stress (Hamerlynck et al., 2000). Unfortunately, little is known of the effect of water deficit stress on the desert plant species Calotropis procera. Generally, the effect of environmental stress on the photosynthetic machinery was not investigated except the studies carried out in our laboratory at Taibah University (Al-Madinah Al-Munawwarah, Kingdom of Saudi Arabia) (Akhkha, 2009 in press). In this study, photosynthesis rate was not affected at low water regime and even increased at mild water regime. How such effect is reflected on chlorophyll fluorescence is the subject of the present study.
The effects of different water regimes on chlorophyll fluorescence parameters and chlorophyll content were determined in Calotropis procera, which is well adapted to arid regions environment such as AlMadinah Almunawwarah. As far as we know, the present work is the 1st to use chlorophyll fluorescence technique to study the effect of different water regimes on the photosynthetic machinery of C. procera.
Materials and Methods
Plant Material & Growth Conditions
Mature fruits of Calotropis procera were collected from plants grown in the region of AlMadinah AlMunawwarah, Kingdom of Saudi Arabia. The fruits were placed in paper envelops and left to dry under room temperature conditions for 2 weeks. Seeds were collected, washed and then soaked in distilled water for 24 hours prior to germination. Seeds were placed on distilled water saturated Whatman No 2 filter paper in Petri dishes and then incubated in the dark at 30ºC ± 2ºC. Seedlings of uniform size were planted in plastic pots filled with compost and sand mixture (3:1). The pots were placed in a growth cabinet at a temperature 28ºC ± 2ºC. Illumination was provided by fluorescent tubes lamps giving 150 µmol quanta m-2 s-1 at plant level during 16 hours photoperiod. The relative humidity within the cabinet was around 40% ± 5%.
Chlorophyll Fluorescence Measurements
Chlorophyll fluorescence parameters were measured at the fourth youngest leaf of four weeks old Calotropis with a Hanastech Chlorophyll Fluorometer (Hansatech Instruments, Narborough Road, Pentney, King’s Lynn, Norfolk, England PE32 1JL, UK). A leafclip was attached to a leaf and the shutter plate was closed to start the dark adaptation, making sure that the leafclip is not directly exposed to the light source as light spoils the dark adaptation. When the leaf was dark adapted for 15 minutes, the sensor head was carefully fit over the location ring of the leafclip in order to seal out the light. The shutter plate was opened and the measurement starts by pressing the button on the sensor head. The chlorophyll fluorescence parameters are recorded for each fourth leaf of each plant.
Chlorophyll Levels Determination
Chlorophyll levels in the fourth leaf of Calotropis plants were determined as Chlorophyll Index using a non-destructive method that uses an optical instrument called Chlorophyll Meter (Apogee Instruments Inc., 721W 1800N, Logan, UT 84321, USA).
Data Analyses
Analysis of variance (ANOVA) was performed on all the data using Minitab version 11.0. The values presented are the means of six replicates ± S.E.
Results and Discussion
Chlorophyll fluorescence parameters such as initial fluorescence Fo, maximum fluorescence Fm, maximum primary yield of photochemistry of photosystem II (Fv / Fo), maximum quantum yield of photosystem II (Fv / Fm), were measured in the forth leaf of plants treated with one of the three water regimes (30%, 50% and 80%). The results are summarised and plotted graphically in Figs 1, 2, 3 & 4.
Effect of water stress on initial fluorescence Fo
Initial fluorescence (Fo) is fluorescence level when plastoquinone electron acceptor pool (QA) is fully oxidized (Rong-Hua et al., 2006). Fo is affected by any environmental stress, which causes alterations at the pigment level of PS II such as thermal stress (Reigosa Roger & Weiss, 2003). The present study shows that Fo was decreased (Fig. 1) due to water deficit stress in both water regimes 30% and 50% compared to the control (80%). However, the decrease was only significant (p < 0.05) at 50% water regime at the second harvest. Such decrease in Fo was interpreted by the possibility that water deficit stress caused some damage to the regulatory processes external to the reaction center of PSII (P680). According to Hong and Xu (1999), the decrease of Fo reflects the destruction of photoprotective processes that helps the dissipation of excess energy with the leaf.
However, leaves under 30% water regime showed slight recovery at the 3rd harvest to a level even higher than that of the control leaves but statistical analysis showed that differences were not significant (p > 0.05). This indicates that water deficit did not affect significantly the integrity of the light harvesting complex on the PS II.
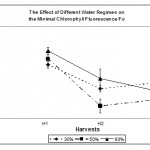 |
Figure 1
|
Effect of water stress on maximal fluorescence Fm
The fluorescence rises from Fo when the primary acceptor QA is oxidised to maximal fluorescence Fm when all reaction centers become closed (Ranalli et al., 1997). Fig. 2 shows that Fm was significantly (p < 0.05) lowered by both low and mild water regimes 30% and 50% respectively in the first harvest indicating an inhibition of PS II. Later on, Fm was increased in the 50% water regime to a level similar to that of the control indicating a sort of reversible recovery of the PS II from being inhibited. In the 30% water regime, Fm not only recovered but reached levels higher than that of the control; however, the differences were not significant (p > 0.05). At the third harvest, non of the differences in Fm between the difference water regimes were significant (p > 0.05). This suggests that, on the onset of water deficit stress at both low and mild water regimes, PS II was inhibited. However, leaves were able to recover at the second and third harvests demonstrating that C. procera leaves were able to recover from the inhibition of PS II. How such recovery takes place is not known and not investigated in the present study. We could conclude that C. procera leaves were able to adjust the level at which the electron acceptor QA is saturated to cope with water deficit stress.
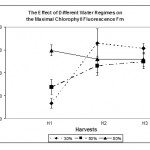 |
Figure 2
|
Effect of water stress on Photochemistry activity
The other parameters such as Fv, Fv / Fm, Fv / Fo characterise the functional state of PS II in dark-adapted leaves (Vassilev & Manolov, 1999).
Maximum/potential quantum efficiency of PS II (Fv / Fm)
The Fv/Fm ratio is considered to be a measure of PSII effectiveness in the primary photochemical reactions (Buttler, 1977). It is proportional to the quantum yield of the photochemical phase of the photosynthesis and it is used thoroughly as an indicator of stress induced on the photosynthetic apparatus (Reigosa Roger & Weiss, 2003).
In the present study, Fig. 3 shows that Fv / Fm ratio was reduced significantly in both mild and low water regimes 50% and 30% in the first harvest. According to (Rong-Hua et al., 2006), decrease in Fv / Fm ratio indicates that PS II is damaged and the primary reaction of photosynthesis may be inhibited. (Reigosa Roger & Weiss, 2003) stated that a decrease in such ratio is a good indicator for damage of photoinhibition caused by the light when the leaves are under environmental stress such as water deficit, cold, freezing, excessive illumination or high salinity.
At The second and third harvests, the ratio increased to levels higher than that of the control plants (80% water regime). However, such increase was not statistically significant (p > 0.05). This indicates that the effectiveness of the PS II of stressed leaves is back to normal if it is not even enhanced. We suggest that any damage happening at the onset of water deficit stress was reversed and the PS II activity of stressed leaves was recovered, indicating a high adaptability of Calotropis leaves to water deficit stress.
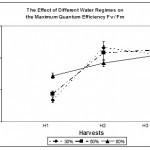 |
Figure 3
|
Maximum primary yield of photochemistry of PS II (Fv / Fo)
The maximum primary yield of photochemistry of PS II (Fv / Fo) provides an estimation of leaf photosynthetic capacity (Rong-Hua et al., 2006). Similarly to Fv / Fm, Fv / Fo ratio (Fig. 4) was significantly (p < 0.05) decreased at the first harvest in both mild and low water regimes which indicates that PS II was damaged (Fig. 4). However, such damage was reversible as the ratio increased significantly (p < 0.05) again to levels higher to that of the control leaves. The third harvest showed no difference in the ratio between all water treatments. The decrease in Fv / Fo was observed by other authors in different barley genotypes (Rong-hua et al., 2006).
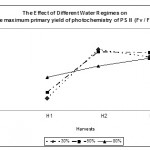 |
Figure 4
|
Chlorophyll Content
Chlorophyll content was measured as Chlorophyll Index using the Chlorophyll Meter (see Materials & Methods). Fig. 5, shows that extreme and mild water regimes 30% and 50% respectively had a highly significant (p < 0.01) effect on chlorophyll content measured as chlorophyll index. Both regimes increased the chlorophyll content in the leaves in order to cope with the water deficit stress and to compensate for any effect that could affect the function of the photosynthetic machinery. This phenomena act as a defence mechanism against abiotic stresses that can be detrimental to the plants. How this desert species preserve the photosynthetic machinery and tolerate water deficit stress is not known and more work has to be carried out to understand such mechanism(s) in order to use it for future crop plants to withstand shortage of water especially in arid regions such as the Kingdom of Saudi Arabia. In other studies, the level of chlorophyll and chlorophyll a + b ratio were not affected by water deficit (Munné-Bosch et al., 2006). In contrast, Guerfel et al. (2009) reported that water deficit decreased the level of chlorophyll (a + b) content in different Tunisian olive varieties (Olea europaea L.). The decrease in chlorophyll due to water deficit was attributed to the sensitivity of this pigment to such stress, which has been reported by several researchers (Younis et al., 2000; Moran et al., 1994). In the present study, Calotropis plants showed high insensitivity of the chlorophyll pigment to water stress suggesting certain mechanism(s) protecting the photosynthetic machinery from such stress. We conclude that Calotropis procera plants are highly tolerant to water deficit stress which explains the insensitivity of its growth in very harsh environments such as drought, which makes it a very successful species in arid regions such as Saudi Arabia.
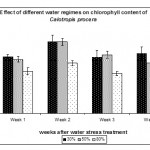 |
Figure 5
|
Chlorophyll Fluorescence versus Chlorophyll Content
The relationships between Chlorophyll Index and some fluorescence parameters and ratios were plotted in Figs. 6, 7, 8 & 9.
Chlorophyll Index was found to be positively correlated with Fo in both stressed and non-stressed plants (Fig. 6). Such correlation was also observed in barley plants under stress. However, well watered barley plants showed insignificant (p > 0.05) negative correlation between Chlorophyll Index and Fo (Rong-hua et al., 2006).
Chlorophyll Index was found to be positively correlated with Fm when plants are well watered (Fig. 7). However, when plants were subjected to water deficit stress Chlorophyll Index was negatively correlated with Fm at both 30% and 50% water regimes.
Similarly, Chlorophyll Index was found to be strongly negatively correlated with Fv / Fo ratio in the two water regimes 30% and 50% (Fig. 8). Well watered plants showed negative but lower correlation compared to that of low and mild water regimes. However, no significant (p > 0.05) correlation was observed between Chlorophyll Index and (Fv / Fm) ratio in any of the water regimes (Fig. 9). In contrast, Rong-hua et al. (2006) showed that well watered plants showed no significant correlation between chlorophyll content and any of the fluorescence ratios. However, a strong positive correlation was observed under water deficit conditions.
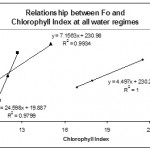 |
Figure 6
|
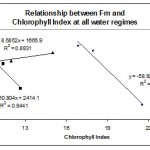 |
Figure 7
|
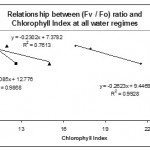 |
Figure 8
|
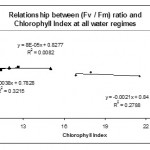 |
Figure 9
|
Conclusion
In the present study, we conclude that the desert plant species Calotropis procera was affected by limited water deficit regimes on the onset of the water stress. As the stress continues, the photosynthetic machinery started recovering and the effectiveness became similar or even better than that of the well-watered plants. This indicates that any damage caused to the photosynthetic machinery on the onset of water stress, was reversible and plants became well adapted to water limitations. Whether this recovery was due to the increase on chlorophyll content and / or increase in the effectiveness of the photosystem II light harvesting complex needs more investigation and future work in our laboratory at Taibah University (AlMadinah AlMunawwarah, Kingdom of Saudi Arabia) will reveal such mechanism(s).
References
- Akhkha, A. 2009. The effect of water stress on photosynthesis, respiration and relative chlorophyll index of the desert plant Calotropis procera. Biosciences, Biotechnology Research Asia, 6(2), in press.
- Araus, J.L. & Hogan, K.P. 1994. Comparative leaf structure and patterns of photoinhibition of the neotropical palms., Scheelea zonensis and Socratea durissima growing in clearing and forest understory during the dry season in Panama. American Journal of Botany, 81: 726-738.
- Araus, J.L., Amaro, T., Voltas, J., Nakkoul, H. & Nachit, M.M. 1998. Chlorophyll fluorescence as a selection criterion for grain yield in durum wheat under Mediterranean conditions. Field Crops Research, 55: 209-223.
- Buttler, W. L., 1977. Chlorophyll fluorescence: a probe for electron transfer and energy transfer. In: Encyclopedia of Plant Physiology, 5: 149–167.
- Guerfel, M., Baccouri, O., Boujnah, D., Chaïbi, W. and Zarrouk, M. 2009. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Scientia Horticulturae, 119(3): 257-263.
- Hamerlynck, E. P., Huxman, T. E., Loik, M. E. and Smith, S. D. 2000. Effects of extreme high temperature, drought and elevated CO2 on photosynthesis of the Mojave Desert evergreen shrub, Larrea tridentate. Plant Ecology, 148(2): 183-193.
- Havaux, M. and Lannoye, R. 1983. Chlorophyll fluorescence induction: A sensitive indicator of water stress in maize plants. Irrigation science, 4(2): 147-151.
- Hong, S.S. & Xu, D.Q. 1999. Light-induced increase in chlorophyll fluorescence Fo level and the reversible inactivation of PS II reaction centers in Soybean leaves. Photosynthesis Research, 61: 269-280.
- HongBo, S., ZongSuo, L., MingAn, S., ShiMeng, S., and ZanMin, H. 2005. Investigation on dynamic changes of photosynthetic characteristics of 10 wheat (Triticum aestivum L.) genotypes during two vegetative-growth stages at water deficits. Colloids and Surfaces B: Biointerfaces, 43: 221-227.
- Ingram, J., Bartels, D., 1996. The molecular basis of dehydration tolerance in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 47: 377-403.
- Massacci, A., Nabiev, S.M., Pietrosanti, L., Nematov, S.K., Chernikova, T.N. & Leipner, J. 2008. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under filed conditions studied by Gas-Exchange analysis and chlorophyll fluorescence imaging. Plant Physiology & Biochemistry, 46(2): 189-195.
- Moran, M., Becan, I. Iturbe-Ormaetyl, S. Frechilla, R.V. Klucas and Aparicio-Tejo, P. 1994. Drought induces oxidative stresses in pea plants. Planta, 194: 346–352.
- Munné-Bosch, S. & Cela, J. 2006. Effects of water deficit on photosystem II photochemistry and photoprotection during acclimation of lyreleaf sage (Salvia lyrata L.) plants to high light. Journal of Photochemistry and Photobiology, 85(3): 191-197.
- Ranalli, P.M., Candilo, D. & Bagatta, M. 1997. Drought tolerance screening for potato improvement. Plant Breeding, 116: 290-292.
- Reigosa Roger, M. J. & Weiss, O. 2003. Fluorescence Techniques. In: Handbook of Plant Ecophysiology Techniques. Ed. By: Reigosa Roger, M. J. Kluwer Academic Publishers. New York.
- Rong-hua, L., Pei-guo, G., Baum, M., Grando, S. & Ceccarelli, S. 2006. Evaluation of chlorophyll content and fluorescence parameters indicators of drought tolerance in barley. Agricultural Sciences in China, 5(10): 751-757.
- Souza, R.P., Machado, E.C., Silva, J.A.B., Logôa, A.M.M.A. & Silveira, J.A.G. 2004. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in Cowpea (Vigna unguiculata) during water stress and recovery. Environmental and Experimental Botany, 51: 45-56.
- Vassilev, A. & Manolov, P. 1999. Chlorophyll Fluorescence of barley (Hordeum vulgare L.) seedlings grown in excess of Cd. Bulg. J. Plant Physiol., 25(3–4): 67–76.
- Younis, M.E., El-Shahaby, O.A., Abo-Hamed, S.A. & Ibrahim, A.H. 2000. Effects of water stress on growth, pigments and 14CO2 assimilation in three sorghum cultivars. Agron. Crop Sci., 185(2): 73–82.

This work is licensed under a Creative Commons Attribution 4.0 International License.



